ヒマラヤ高山帯の草地群落と放牧の影響 |
|
高槻 成紀 東京大学総合研究博物館 |
ヒマラヤの高山帯の植生配列はいくつかの環境要因によって支配されている[菊池一九九二]。植物群落の配置は、群落を構成する植物の生理的特性に応じて適地が決まっており、それが例えば地面の温度であったり、湿度であったりするために、標高や斜面の向きなどによって出現する植物群落に違いがある。ヒマラヤの高山帯のような極限の環境にあっては、環境の違いが明瞭であるため、植物群落の配列は温暖で湿潤な地域とは比べものにならないほど明確な違いが現れる。ヒマラヤが環境と群落の成立という生態学的なテーマを考えてゆく上でまことに好適な場所といわれる所以である。 ところで私たちは自然な環境といえば人工的な環境、例えば都市とは対照的なところを想定する。都市よりも田園地帯に自然があり、田園地帯よりも山村に自然がある。日本では海岸に町があり、平野に水田があり、丘陵地に里山があり、山に入ると山村や森林がある。日本だけでなく世界のどこでもおよそそのような構図がある。そうであれば、「世界の屋根」と呼ばれるヒマラヤはまさに最もよく自然が残された場所であると考えるのはごく当然なことであろう。確かに七〇〇〇メートルにも達する氷雪の世界には人跡未踏の地が多く残されており、現在でもそれを目指して多くの登山隊が競い合っている。しかしそのすぐ下(といっても富士山などよりははるかに高いのだが)に拡がる植物の世界はどうだろうか? そこが植物にとって極限の環境であり、環境要因の違いによって植物群落が明瞭に住み分けているのは前述の通りである。私はそのことを踏まえた上で、ヒマラヤ高山帯の植物群落をもうひとつ違う視点から眺めてみたいと思う。
ネパール中部の湿潤地域では標高およそ三〇〇〇メートルで森林限界となる。日本などと違って落葉広葉樹林は未発達だから、常緑広葉樹林がそのまま森林限界になることもあれば、モミ属やトウヒ属などの針葉樹林に移行してから森林限界になることもある。いずれにしても森林限界を越えると、そこは広々とした景色となる[挿図1]。山を登ってゆくと、初めのうちは森林が断片的に残っていたり、低木群落が多かったりするが、三五〇〇メートルを越えると、しだいに草本群落が多くなってくる。一方、乾燥地域であるパキスタン北東部のカシミールでは低地は砂漠に近い、植物の乏しい世界であり、逆に三五〇〇メートル以上になって植物が豊富になった。これは高地に雪が積もって、夏の間その融水が供給されるためである[挿図2]。いずれの地域でも高山帯は放牧に適した草地が発達していた。
草地はイネ科が優占していることが多い。そしてそのような群落でのどかに草を食むヤギやヒツジ、それにウシまたはウシとヤクの雑種などの家畜の姿をよく見る[挿図3]。また、ネパールも中部以西ではウマやロバも多い[挿図4]。歩いているといたるところに家畜の糞が落ちている。ときどき通り過ぎる集落では家畜が特に多くなり、そういうところでは家畜が入らないように耕作地を石垣で守っている。四〇〇〇メートル以上になると定住する集落はなくなり、わずかに石造りの粗末な小屋があるだけになる。これは夏の放牧のために牧民が寝泊まりに使うためのもので、カルカと呼ばれる。カシミールを訪れたときはところどころにこのような家畜を集める場所があり、その周辺ではおびただしい数の糞が積み重なるように積もっていた。
ここで草食獣による採食と草本群落について簡単に触れておきたい。ススキ群落は日本のいたるところで見られる代表的な草本群落であるが、これは放置すると次第に低木群落となりやがては森林群落に移行する。一方、適当な密度の家畜を放牧するとススキ群落のまま維持されるが、放牧圧が強いと背丈の低いシバ群落へと移行する。ススキ群落のような群落を長草型群落、シバ群落のような群落を短草型群落という。したがって短草型群落があれば、相当強い放牧の影響があると判断でき、その意味で放牧圧の指標となる。 短草型群落と長草型群落とは、草丈が高いか低いかという構造的な違いだけでなく、植物のもつもう少し生物学的な特性を反映している。イネ科植物の構造はいずれの種でも基本的に似ており、ストローのような茎(稈(かん))に平行脈の走る細長い葉がついている。よく見ると、稈全体はソフトクリームのコーン(それよりはよほど細長いが)を重ねたような構造になっており、コーンの先端に葉がついている。これらのコーンが伸び切ると、内側から新しいコーンを伸ばして、草丈全体が高くなってゆく。これらのコーンは節にある生長点から伸びてくる。ススキは草丈二メートルもあり、シバは一〇センチにも満たないが、このような基本的な構造はススキでもシバでも変わりはない。要するにシバはススキをうんとコンパクトに縮めた形をとっているとみることができる。 このことを草食獣の採食と合わせて考えてみたい。草食獣は葉を食べる。ススキのような長草だと葉とともに稈を食べるから生長点も奪われることになり、植物にとってはダメージが大きい。それでも双子葉植物のように生長点が茎の先端にあるのに比べれば、最下部の節が残っていればそこからある程度の再生が可能である。採草によってススキ群落が維持されるのはこのようなススキの特性による。これに対してシバの場合、節はほとんど地上ギリギリにある。ていねいに草を食べるヤギのような家畜でも、シバのように背の低い草を食べる場合、葉の一部を食べるだけで生長点は残してしまう。そのため採食はシバにとってダメージにならない。このような違いのために採食圧が強くなるとシバが有利になり、次第にススキを凌ぐようになる。 以上はススキとシバの比較であるが、これは基本的にそのほかの長草型群落と短草型群落にもあてはまる。前述の説明では短草型群落の強さを強調したが、それは採食という、一種の特殊な環境要因が強くかかる条件下での強さであることに注意する必要がある。短草型群落を構成する植物は、その特殊な影響がない場合はむしろ弱い植物であることが多い。その最大の弱点は耐陰性である。一般に、短草型群落の構成種は採食に耐性がある一方で、暗い環境では生育しにくいという弱点をもっている。採食は、そのほかの採食に耐性のない植物を除去するという効果をもつから、この点で草丈の低い植物に有利となる。植物体の一部を除去する採食や、踏み付けなどの厳しいストレスは、普通の植物にとってはマイナス効果をもつが、光をめぐる熾烈な種間競争においては草丈の低い植物にはまたとない好条件を提供する。これが植物群落にとっての採食のもつ最も重要な意義である。
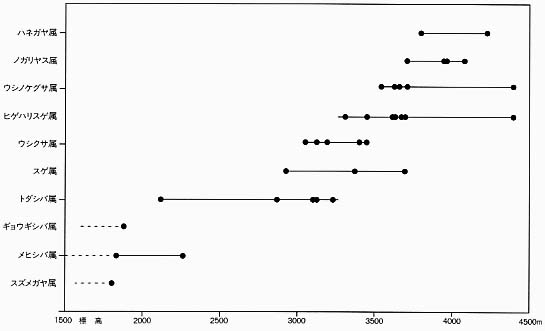
さて、そのような視点からヒマラヤ高山帯の植物群落をながめるとどうなるだろうか。ネパールの三地域での調査ではほとんどの場所で短草型群落がいたるところで見られた。それどころか、比較調査をしようとして長草型群落を探しても、なかなか見つからなくて困るというのが実情だった。短草型群落の優占種は標高によって違い、ネパール中部の高山帯ではトダシバ属などが低い標高に出現し、三〇〇〇メートルを越えるとウシクサ属、ヒゲハリスゲ属、三五〇〇メートル以上ではウシノケグサ属、ノガリヤス属などへ移行していった[挿図5、高槻一九九三]。
最も注目されるのはヒゲハリスゲ属で、短草型群落の優占種の中でこれだけがイネ科ではなく、カヤツリグサ科であった。日本語では区別しないが、「草」には草本を意味するherbと、イネ科を意味するgrassとがある。前述の短草型群落はshort・grass typeであり、厳密な意味ではヒゲハリスゲ属の優占する群落はgrasslandではないことになる。sedge・landというべきかもしれないが、イネ科、カヤツリグサ科、イグサ科をまとめてgraminoidsあるいはgrasslike plantsとしてまとめることも多いから、広義のgrasslandの意味で短草型群落としてよいだろう。このヒゲハリスゲ属群落も家畜によってよく利用されており、その意味でこの地域で重要な資源となっていた[挿図6]。
ところで、ヒゲハリスゲ属はスゲ属にはなはだよく似た植物で、日本では北海道と本州中部の高山帯に一種だけが生育しているにすぎない。しかし中部ネパールの標高三〇〇〇メートル以上では面積的にも量的にもほかのイネ科を凌駕しているばかりでなく、種数も豊富で、このグループがヒマラヤの高山帯で種分化をとげるとともに重要な植生の構成要素であることがわかった。ヒゲハリスゲ属の分類・系統と生態に関してはユーラシア全体を視野に入れた総合的な研究が必要であろう。
これら短草型群落の特徴を家畜の採食を考えながら検討してみよう。実際にこれらの群落が家畜に利用されていることは、例えば構成種の多くに家畜の食べ跡があること、この群落に糞や足跡が多いこと、家畜の糞を分析してみた結果、短草型群落の主要構成種が多く出現したことなどから明らかである。 群落構造という点からいえば、短草型群落は構造が単純であるということで特徴づけられる。植物群落は通常、垂直的にいくつかの階層構造をもつ。森林群落では最低でも三層あり、長草型群落は上層と地表を被う二層からなる。後者の場合、上層には優占種である丈の高い草とそれにからまるツル植物などが、下層には丈の低い植物しか生育しない。この点、短草型群落はこの下層に相当する層だけしかなく、構造的には極端に単純である[挿図7]。この層で生育可能な植物は生育型でいえば、匍匐型、ロゼット型などが主体となる。優占種の多くはイネ科とヒゲハリスゲ属であったが、これらはシバ同様次々と葉を展開する「採食適応型」で、生育型は匍匐型が多かった。また、オオバコ属やセリ科のいくつかの種のように、ロゼット型で家畜の踏み付けに耐性のある種も多かった。これとは別にツメクサ属やスズメノヤリ属などのように、小型であることによって、これら採食に耐性のある植物の間にまぎれこむように生育している植物もあった。
短草型群落のひとつの特徴は限られた種(しばしば一種)が飛び抜けて優占することにある[高槻一九九三]。その理由は明らかでないが、生育型にひとつのカギがありそうなので、これに注目して調査を続けている。一方、このことは、採食が群落の多様性を低下しているということを示しており、保全生物学的にも重要な意味をもつものと考えられる。
ところで、ここまで短草型群落にしぼって紹介してきた。確かに短草型群落は草食獣による採食影響と最も深く結びついた群落ということができる。その要点は採食されることによって自らの存在を確保しているということにある。繰り返しになるが、採食は植物体を除去することであるから、生理学的にいえば疑いなく植物にとって有害なことである。それでも採食が短草型群落構成種にとって有利になるのは、植物には光をめぐってつねに他種との競争があり、採食によってその競争に勝てるという一事があるからである。彼らの採用した対採食戦略は、あえて採食されるという危険をおかすことによって自らを有利にするという逆説的なものである。しかし採食に対する植物の戦略にはもっと単純なものもある。それは食べられることに対して食べられなくするというものであり、その意味においてこれは「防衛」と呼ばれる[高槻一九八九]。これには大きく分けて二つあり、ひとつは化学防衛、もうひとつは物理防衛である。化学防衛とは植物体内に特殊な物質を含むことにより動物に食べられなくするもので、有毒植物や不快な匂いのするさまざまな植物が知られている。一方、物理防衛とは植物体を堅くするなどの方法で食べられなくするもので、トゲがその代表である[高槻一九九六]。 これら採食防衛をする植物をヒマラヤで観察したところ、いくつかの例が見られた[高槻一九九三]。化学防衛植物は量的には少なかったが、シャクナゲ属、サクラソウ属[挿図8]、アヤメ属、キジムシロ属[挿図9]などが目立った。一方、トゲ植物は多く、ムレスズメ属[挿図10]、ゲンゲ属などマメ科が多いのがひとつの特徴であったが、そのほかにもノイバラ属、メギ属、アザミ属、イラクサ属などもあった[高槻一九九三]。挿図11のモリーナ・ポロフィラもその例で、植物体全体がトゲのかたまりのような植物であった。ネパールのランタン谷の標高三五〇〇メートル付近のある低木群落は群落構成種の八〇パーセントほどがトゲ植物であり、ウシなどは食べないで通り過ぎていた。もっともヤギはトゲにもめげずそれらの葉を細かい口先で摘み取って食べていたが、それでも普通の植物のように枝ごと折り取ることはないので、トゲは明らかに防衛の機能を果たしていた。
このような結果、採食圧の強い場所では短草型群落と防衛植物だけが目立つ、特異な景観を呈するようになる[挿図12]。
さて、私は本文を都会の対極としての秘境ヒマラヤから始めた。それは動植物や人間にとって極限の環境であり、人々は人跡未踏の無垢の自然があるものと期待する。しかし実際のヒマラヤの高山帯は、少なくとも植物群落の特徴から見ると、いたるところが家畜の強い採食の影響を受けているというのが実態である。私がみた範囲はごく限られるものの、これはこれまでヒマラヤを訪れた多くの植物学者が共通して指摘するところである。その意味ではヒマラヤの動植物は決して「自然」ではない。美しい高山植物が咲き乱れるお花畑も、実は家畜の採食影響下で維持された短草型群落であることが多い。それらの群落は低温と短い夏に制約されたフローラが地形に基づいて群落配列をしている。こうして成立した植物群落が毎日毎日、家畜の絶え間ない採食にさらされて、あるいは変形し、あるいは種組成を変えながら存続している。これが「秘境ヒマラヤ」の植生の実態であるらしい。そのことを踏まえた上で生物の保全という観点から三つのことを指摘したい。 ひとつは、それではヒマラヤにおけるこのような人為影響はいつから始まったのかという点である。花粉分析の情報によると、ナラ属が減少し、マツ属などが増加し、人為影響が始まったことを示唆する年代は控えめにみても二〇〇〇年を下ることはない[三好・星野一九九七]。そうであるとすると、ヒマラヤ高山帯の植生はごく短い間に急激な変化をしたことになる。改めて思うことは、人間による自然への働きかけが激しくなったのは生物の歴史からすればごく最近のことであり、しかもそれが突然に大規模かつ強度なものになったということである。ヒマラヤ高山帯の群落研究は、その渦の中に大ヒマラヤさえ飲み込まれていたことを示唆している。そのことは、私たちに生物の保全もまたそれだけ緊急かつ大規模でなければならないことを改めて警告している。 もうひとつの問題は、このような人間による一種の自然破壊は、経済発展を遂げた先進国で進められている自然保護とは全く質が違うものであるということである。先進国ではかつてさかんに自然破壊が行われ、それだけによったのではないにせよ、経済的に豊かになり、いわばゆとりがもてるようになってから自然保護の思想が芽生え、社会運動として定着してきた。しかしヒマラヤの主要部を占めるネパールは世界でも最貧国のひとつである。人々は最低限の生活に耐えながら、少しでも豊かな生活を求めて高山帯まで出かけて放牧をしている。あるいは燃料を求めて高山帯近くまで登ってくる。機械力による森林伐採や工場・自動車による排気ガスを抑制するという形での自然保護とは、規模においても、形態においても、また人間活動と自然とのかかわりという点においても大きく性格が異なる。これを先進国型の自然保護運動と同じ論理で批判することはとうていできないところに問題の難しさがある。 最後の、そしてより一般的な問題としては、ヒマラヤの高地でさえこれだけの人手が入っていることを考えれば、もはや地球上に手つかずの自然はないという事実そのものを認識しなければならないという点である。しかし、そうだからそのような自然が価値がないということにはならない。私たちはそのことを認識した上で、人為影響下の動植物の保全を考えてゆかなければならない。ヒマラヤ高山帯についていえば、現在の植生が採食影響下に成立しているとすれば、採食によって減少した植物があるはずだし、採食のために絶滅した植物さえあるかもしれない。少なくとも採食によって群落の種組成の多様性が低下していることは間違いないと思われる。この地域の保全にあたってはこのような認識に立つ必要がある。これに関連して、ヒマラヤ高山帯は実は野生有蹄類の有力な生息地であったということも忘れてはならない。現在でも北米のビッグホーンに近縁なアルガリや、ヒマラヤタール、ゴーラルなどの有蹄類が生息している。これら多くの種は分布域が狭められ、個体数も減少している。その中には狩猟という直接的な脅威によって絶滅に瀕しているものもあるが、多くの種は家畜の放牧によって生息地や食物を奪われるという、真綿で首を絞めるような間接的な人為影響によって危機に瀕しているのである。もちろん放牧は草食性の昆虫などに様々な影響を及ぼしていることは疑う余地がない。このように、放牧のもつ意味は単に植生への影響というだけでなく、ヒマラヤの生態系全体への影響という視座で捉えなければならない。 |











【参考文献】菊池多賀夫、一九九二年、「ヒマラヤ高山帯の植生とその分布パターン」『遺伝』第四六巻九号、一七−二二頁高槻成紀、一九八九年、「植物および群落に及ぼすシカの影響」『日本生態学会誌』第三九巻、六七−八〇頁 高槻成紀、一九九三年、「ネパール中部、ランタン谷の草地群落と家畜の影響」『プランタ』第二六巻、一〇−一九頁 高槻成紀、一九九六年、「とげの収支決算」、週刊朝日百科『植物の世界』第九四巻、三一八−三二〇頁 Miyoshi, N. and Hoshino, T. 1997. Pollen analysis of Quaternary sediments taken from Jomosom in Annapurna Himal Region, Nepal. In: T.Hoshino ed., Biosystematic studies of Gramineae and Cyperaceae in Nepal pp.68-74. Okayama University of Science. |
| 前頁へ | 表紙に戻る | 次頁へ |