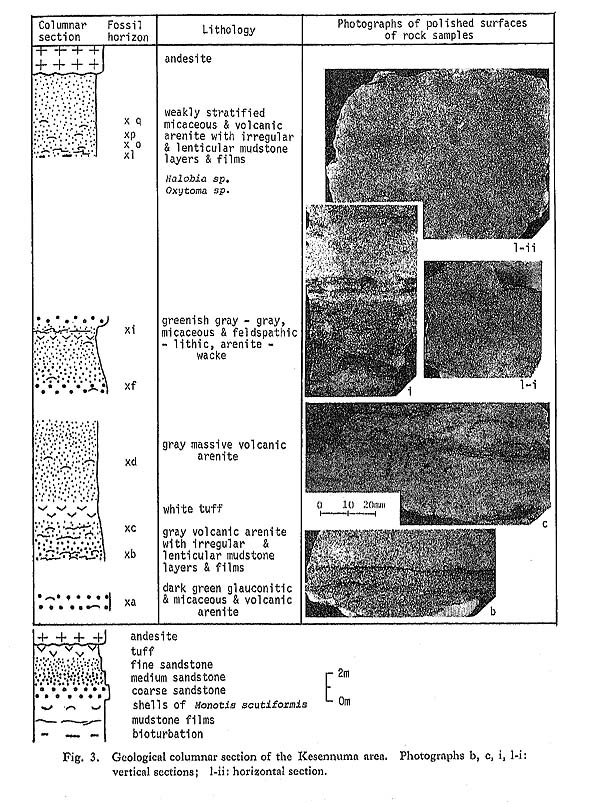
CHAPTER 6
Chronological Changes of Shell Morphology and Inferred Phylogeny of Monotis
|
Evolutionary changes in a widely distributed taxonomic group should not be generally discussed only on the evidence from local areas. However, several Monotis sequences in Japan, as described below, show essentially the same stratigraphic change in various morphological characters. Although precise correlation among these sequences is still difficult, I believe that the observed morphological shift is mostly evolutionary instead of ecophenotypic. In this chapter morphological changes of the shells with time are described and the evolution of Monotis is reconstructed from the evidence in Japan and the world. 1. Chronological Changes in Some Selected Sequences1) Monotis Sequence in the Utatsu AreaThe chronological changes of shell morphology along the Shirayamazawa section in the Utatsu area are partly demonstrated in Fig. 29 in regard to some numerical parameters. In the two species, M. ochotica and M. zabaikalica, Pp tends to decrease in upward sequence. In other parameters, however, it is difficult to detect any significant changes with certainty.
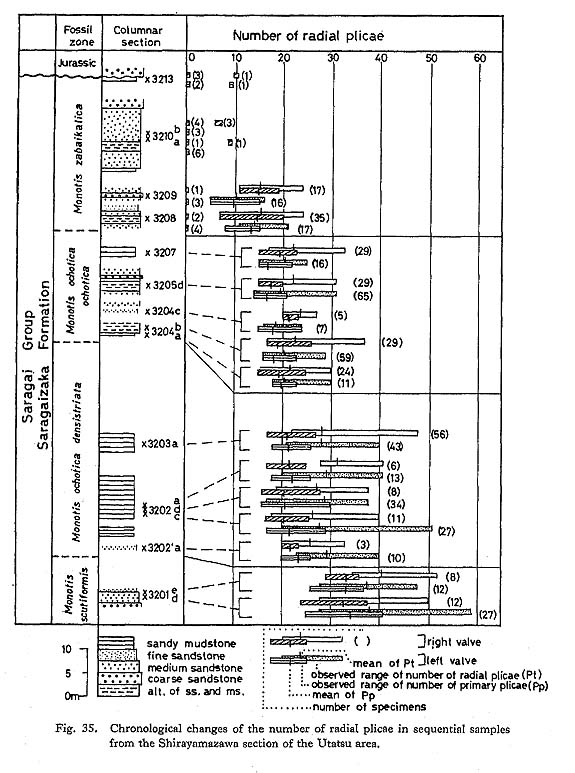
The observed ranges and means of Pt and Pp in each sample are shown together with the stratigraphical columnar section at Shirayamazawa (Fig. 35). The values of Pt and Pp seem to change abruptly at some horizons between 3201e and 3202'a, and between 3207 and 3208, But the change is not so clear at the boundary between the M. ochotica densistriata and M. ochotica ochotica Zones.
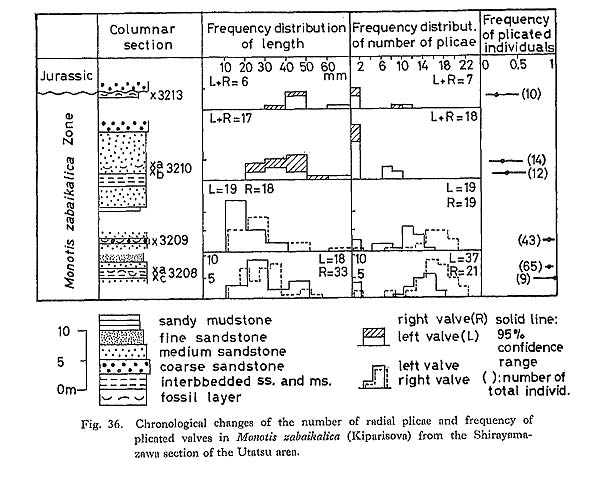
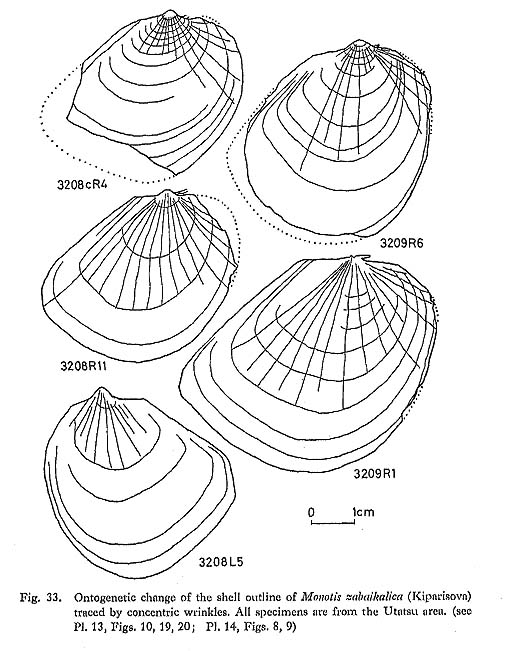
Gradual changes can be recognized within the M. ochotica Zone, namely the mean of Pp gradually changes from 23.1 in sample 3202'a (left plus right valves) to 16.7 and 17.6 respectively in left and right valves of sample 3205d. Sample 3207 has a wider variation in strength of plicae, especially on the posterior wing. On the other hand, M. zabaikalica shows a different mode of chronological change (Fig. 36): Pt and Pp decrease generally with increasing frequency of non-plicated valves. Furthermore, weakening of radial sculpture is also observed throughout the ontogeny of M. zabaikalica (Fig. 33; Pls, 13, 14). In many specimens radial plicae decrease and become obscure with growth. They often disappear simultaneously at a certain concentric wrinkle. In upward sequence, the stage at which radials disappear becomes accelerated. Concentric wrinkles or folds commonly appear instead of radial sculpture, probably for the sake of maintaining shell strength. The stage of the first appearance of concentric wrinkles is also accelerated.
2) Monotis Sequence in the Miyamadani AreaIn the Miyamadani area, M. ochotica densistriata, M. ochotica ochotica and M. mabara in upward sequence show interesting morphological changes (Fig. 21; Pls. 11, 12). The frequency distribution of Pp does not seem to show any significant chronological change from samples a to n. But the earlier samples belonging to M. ochotica densistriata are characterized by more numerous higher-order plicae. M. ochotica ochotica of late stage, which contains abundant "sparsicostata" (Teller (1886)'s variety) type individuals, appears after a barren part between g and i. Toward the horizon n, the frequency of "sparsicostata" type individuals decreases, while that of the "eurhachis" type individuals increases. M. ochotica ochotica of the latest stage is represented by samples o and p, which are composed of individuals with slightly larger numbers of Pp and smaller numbers of higher-order plicae. Values for Pp appear to change rather abruptly between samples p and q. The boundary between the M. ochotica ochotica and M. mabara Zones is placed at this interval. The latest species M. mabara, represented by the latest samples q to w, may indicate that Pp increases with time, judging from the diagrams and the observation of Pp, though the sample size is small.
3) Monotis Sequence in the Arase AreaThree samples (Ar1301a-c) from the Arase area also indicate chronological change (Fig. 30; Pl. 8). The number of Pp slightly decreases in upward sequence, with mean values changing from 13.4 to 11.6 through 12.2 in left valves. It is probably also the case in right valves. Sample Ar1031a consists of valves having only the minimum varia-tion in numbers, width and strength of the plicae. The next sample Ar1301b from 2 meters above Ar1301a contains valves with obscurely plicated posterior part and con-centric wrinkles. The last sample Arl301c from 0.5 meter above 1301b shows more variable plication from so-called "sparsicostata" type to "densistriata" type (Pl. 8, Figs. 1 and 2, and 3, respectively).
2. Inferred Phylogeny of MonotisTaking the above evidence into account, the evolutionary changes of Japanese Monotis can be schematically inferred as shown in Fig. 37. The vertical axis represents relative time (thickness of strata) and the horizontal axis shows approximate observed ranges of Pp and Pt. Before going further, previous works are reviewed here.
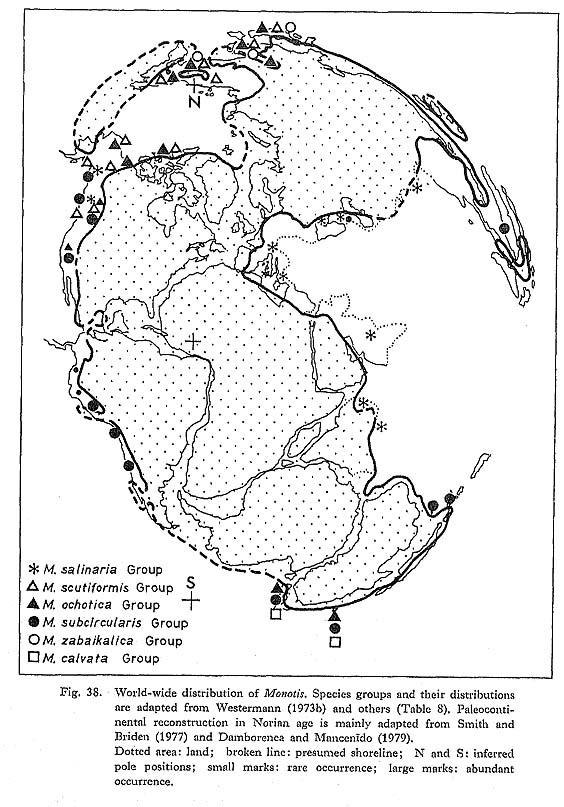
Ichikawa (1954b) recognized the following trends of morphological characters, mainly on the basis of materials from the Utatsu area: Westermann (1973 a, b) suggested the probable phylogeny of Monotis on the basis of such morphological features as convexity and posterior auricles, and proposed a scheme of geographic dispersal in terms of species groups. He detected the trend that equipli-cated species with a weakly plicated and obscurely delimited posterior wing were followed by more variably plicated or smooth and wrinkled species with well developed smooth wings. In other words, the M. scutiformis (=his M. typica) Group was said to have diversified into three species groups, namely the North and Southwest Pacific M. ochotica, Southwest and East Pacific M. subcircularis and Tethyan M. salinaria Groups (Table 8; Fig. 38).
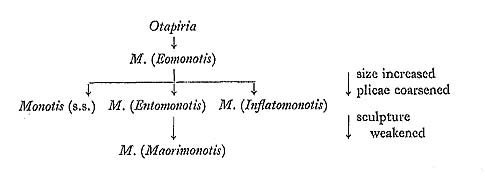
Recently, Grant-Mackie (1980a) commented on the evolution of Monotis (Table 8): "Evolution would have preceded from a small light subequivalved, possibly epi- or pseudo-planktonic ancestor (e.g. Otapiria) to produce firstly Eomonotis, then by size increase and ribs coarsening to Monotis (s.s.) and Entomonotis, followed by weakening of sculpture and reduction of ears (Maorimonotis)". The idea is represented as follows:
Otapiria, which is said to have appeared in Carnian time, before the first appearance of Monotis, is the most closely allied genus. But there is no reliable evidence of the ancestor-descendant relation between the two. Though Maorian Eomonotis does not seem to be the same as the M. scutiformis Group, the first species in the Maorian province is also finely plicated. According to Grant-Mackie, the subgenus might have diversified into three subgenera. Inflatomonotis and Maorimonotis proposed by him are restricted to the Maorian province. The latter, assigned to the M. calvata Group in this study, is an aberrant, non- or weakly-plicated form similar to M. zabaikalica. Let us return to Fig. 37. The first species in the Monotis sequence is M. scutiformis, having a rather small and obliquely elongate outline with numerous fine radial plicae. The later samples probably show slight decrease in Pp and Pt. The second species, M. ochotica shows not only fairly wide intrapopulational variation but also rapid chronological change. The early populations of this species are recognized as a chronological subspecies, M. ochotica densistriata. Because the small specimens of this subspecies, particularly from the lower horizons, are similar to those of M. scuti-formis, it is inferred that M. ochotica was probably derived from M. scutiformis. Within M. ochotica densistriata, Pp decreases gradually and slightly with time. M. ochotica ochotica has fewer, stronger and wider plications than the preceding subspecies. It shows at first rather narrow intrapopulational variation in ornamentation, but the varia-tion later becomes extremely wide. Some individuals have an obscurely plicated but irregularly wrinkled posterior part like M. zabaikalica, and other individuals show fine plication similar to M. mabara. But in the latest stage there is a trend characterized by increase of Pp, in spite of the decrease of higher-order plicae. Among samples from different areas, considerable difference of Pp can be recognized. At present, it is, how-ever, difficult to determine whether this difference is attributed to geographic variation or chronological change. M. zabaikalica and M. mabara coexist in the same fossil beds in the Utatsu, Sakawa, Kubodani and Miyamadani areas, though their relative frequency varies among these areas. While the stratigraphic ranges of the foregoing two species partly overlap, those of M. mabara and M. ochotica ochotica do not. In the serial samples of M. zabaikalica radial sculpture becomes much weaker and concentric wrinkles more prominent with time. The morphological similarity between the young shells of M. zabaikalica and M. ochotica ochotica as well as their stratigraphic relation suggests that the former was derived from the latter. M. mabara is also similar to the densely plicated representatives of M. ochotica ochotica. It is inferred that M. ochotica may be ancestral to M. mabara. Let us consider the phylogenetic pattern of Monotis in Japan and east Siberia. Com-pared with the previous three authors' ideas, the coarsening of plicae and weakening of sculpture are also seen in the Japanese lineage from M. scutiformis to M. zabaikalica. Generally Westermann's (1973b) idea can be accepted, although the M. subcircularis and M. salinaria Groups discussed by him do not occur in Japan. As regards Grant-Mackie's scheme, I think that the phylogeny of Monotis should not be interpreted at the subgeneric level but at species level, because the important unit for evolution is the evolutionary species. The phylogenetic pattern of Japanese Monotis, however, is partly similar to his scheme. Almost all the species of his Maorian Eomonotis, Entomonotis and Inflatomomtis are provisionally included in the M. ochotica Group in this study (Table 8). As in the Japanese lineage, the middle evolutionary stage of the genus seems to be characterized by wide intraspecific variation. The last stage may be represented by the coexistence of the M. calvata and M. ochotica Groups. The chronological change of shell morphology from M. scutiformis to M. ochotica ochotica via M. ochotica densistriata may attribute to phyletic evolution without any speciation, although this represents only one of the lineages of Monotis at that time. But in the last stage of the evolution, the increase of intrapopulational variation might have been followed by the divergence into two lineages, represented by M. zabaikalica and M. mabara. Although the morphology of these two species is predicted by some specimens in the later samples of M. ochotica ochotica, this does not necessarily mean sympatric speciation. In accordance with widely accepted evolutionary theory, repro-ductive isolation between the two lineages seems to have been completed by some allopatric speciation before the first appearance of M. mabara. In the lineage from M. scutiformis to M. ochotica ochotica, the morphological change is regarded as paedomorphic or neotenic, because the morphology of immature growth stages of the ancestral species, especially in shell outline and radial plication, was followed by the mature stage of the descendant species. On the other hand, the phyletic change of ornamentation in M. zabaikalica may be regarded as peramorphic or accelerative, because the disappearance of radial plicae accelerated. McNamara (1982) stated that when "the descendant paedomorphs are suitably adapted to conditions along an environmental gradient, a phylogenetic trend, in the form of paedomorphocline or peramorphocline, can become established." For example, he described the Tegulorhynchia-Notosaria paedomorphocline composed of a series of seven Cenozoic apaedomorph brachiopod species, each of which does not show any directional morphologic shift during its range. In this study no environmental gradient to which different Monotis species were adapting, could be detected. Two morphoclines of Monotis, from M. scutiformis to M. ochotica ochotica, and M. zabaikalica, exhibit the shift within each species. Recently, Williamson (1981) considered that major phenotypic shifts at the Suregei tuff and Guomde levels in late Cenozoic molluscs from the lacustrine Turkana Basin document allopatric speciation, from which the 'punctuated equilibrium' theory could logically result, as proposed by Eldredge and Gould (1972). According to him, an in-crease in phenotypic variance reflecting major developmental instability in transitional populations is followed by reduced levels of variance, comparable with that of typical ancestral populations. The increase in intrapopulational variation at the late stage of M. ochotica ochotica characterized by sample 3207 in the Shirayamazawa section, (Fig. 35), is apparently similar to Williamson's case. The time range of the shift can be cal-culated from 8.3×104 to 1.7×105 years, if the whole Shirayamazawa section of 120 meters had been proportionally formed during 2 ma and if the shift was stratigraphically characterized by a bed 5 to 10 meters thick. Compared with the Williamson's estimation this value is larger, though the difference between the two is not too great. Assuming the same duration of the Shirayamazawa section, the average longevity of Japanese Monotis species can be estimated to be only 0.67 ma. Chronological changes within M. ochotica and M. zabaikalica appear to document the expected result of anagenesis, as recently reviewed by Gingerich (1985). On the other hand, the transition from M. ochotica ochotica to M. zabaikalica and M. mabara may be a result of cladogenesis. |