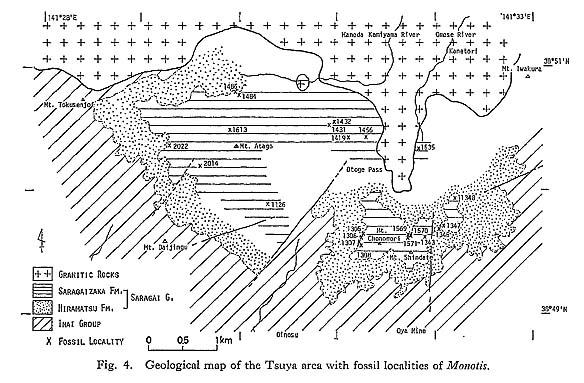
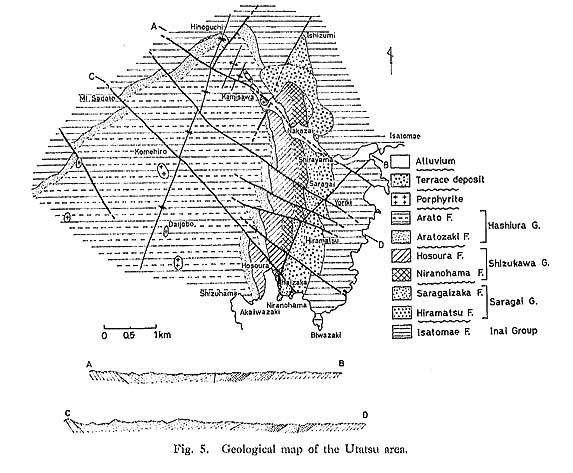
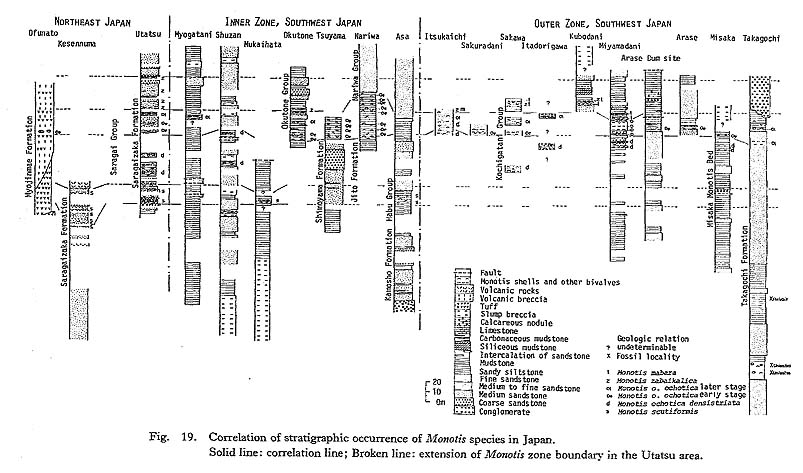
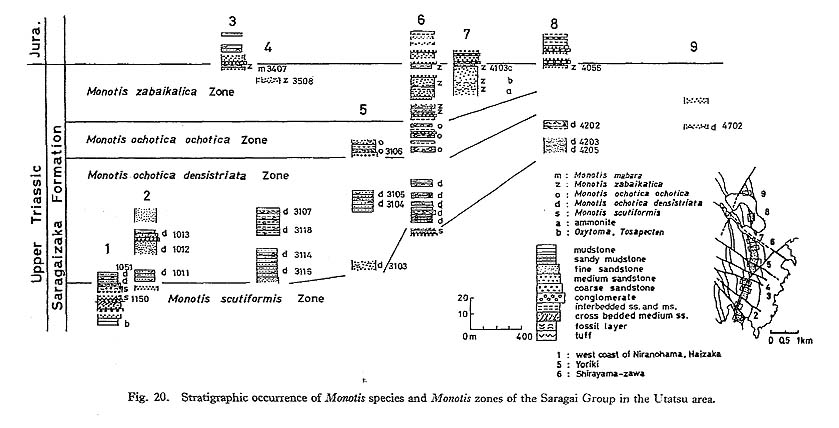
CHAPTER 5
Morphology, Intraspecific Variation, Ontogeny and Relative Growth
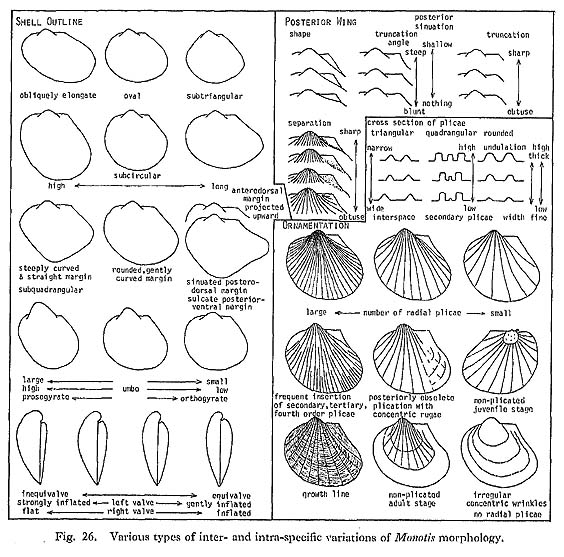
1. Major Morphological Features1) Terms and ParametersDetailed discussions of morphological terms for Monotis valves, were presented by Westermann (1962), Grant-Mackie (1978b) and Payevskaya (1985); I mostly adopt the former two author's terminology here. Figure 25 shows some important morphological terms on the reconstructed valves of M. ochotica ochotica. Various kinds of morphological variations are schematically summarized in Fig. 26.
Monotis is suitable for statistical treatment because of its gregarious and successional stratigraphic occurrence. Intrapopulational and interpopulational morphologic variations can be analyzed by current biometrical methods. The statistical parameters used here shown also in Fig. 25, but their detailed definitions are omitted because they were already well discussed by Westermann and Grant-Mackie. Of various parameters used by the two authors, Pd20, Pd50 and O° are not adopted in the present work, because they are difficult to clarify in the more or less deformed materials. But Ps, P3, P4, Pw and Bl are added. 2) OrnamentationOrnamentation on shell surface of Monotis consists of radial plicae and concentric wrinkles, both of which vary in number, strength and spacing. The mode of radial plication is one of the most familiar and conspicuous characters which many paleontologist have used in the taxonomy. For example, Teller (1886) proposed five varieties of Pseudomonotis ochotica, mainly on the basis of the mode of radial plication. In this species the secondary and tertiary plicae commonly appear with growth at stages of 10-20 and 30-40 millimeters in shell height, respectively. The fourth-order plicae appear only in the latest growth stage. Bifurcation occurs very rarely. Irregular concentric wrinkles or folds are occasionally developed, especially in non-plicated species and on non-plicated umbonal area of some species. The radial plicae of right valve are sometimes more numerous than those of left valves (Figs. 21, 30). Many bivalves have the same number of plicae (or ribs) on opposite valves for interlocking along the plane of commissure. This discrepancy in Monotis is probably due to a difference in the prominence of plicae near the anterior and posterior ends, where the plicae are more or less sharper in the right valve.
3) Byssal Auricle and NotchDetailed observations at the byssal auricle and notch of Monotis were carried out by Marwick (1935), Ichikawa (1958) and Nakazawa (1963), These features are significant in considering the taxonomic position of Monotis. Being very tiny and fragile, however, the byssal auricle can be observed only in well preserved specimens. It is difficult to recognize clear differences in these morphological features among Monotis species. Recently Waller (1984, fig. 1) studied the functional significance of the ctenolium in the Pectinidae. At the same time he made clear that the pseudoctenolium, an analogous (but not homologous) structure to the ctenolium, is often developed in various families of the Pectinacea. According to Waller, the true ctenolium is found only in the Pectinidae and formed as elevations on the surface of an inner foliated calcite layer which is turned outward. The function of the ctenolium, as interpreted by him, is probably to separate the byssal threads by the comb-like teeth and to prevent the body from twisting. The pseudoctenolium is not homologous to the ctenolium, because it is a transformation of the sculpture on the outer surface, even if their function is similar. Some morphological terms concerned with the anterodorsal part of right valve are derived from his definition. Figures 8-11 on Pl. 6, Figs. 4, 5 and 21-23 on Pl. 7 and Fig. 8 on Pl. 9 show the presence of a tiny anterior byssal auricle and pseudoctenolia in some Japanese species of Monotis. The auricle, which is convex outward, projects obliquely upward and inward at first and becomes more parallel to the hinge line in the later stage. It is 0.5-1.5 millimeters wide and 1.5-6 millimeters long (Bl). The byssal fascicle and byssal notch are observed beneath the auricle. A very short byssal fascicle, which is the track of the byssal notch left during growth, has fine growth lines.
Pseudoctenolia can be observed on the lower and upper sides of the byssal notch. Though Marwick did not use the name pseudoctenolium, he already figured the presence of this structure in Monotis (Marwick, 1935, pl. 34, fig. 4; pl. 36, fig. 33). External molds with good preservation from the Nariwa area enable observation of the detailed features. The lower side of the auricle has about 15 transverse ridges, each of which probably has a very tiny spine at the innermost end. The outer five or six ridges, which seem to be an active pseudoctenoliurn projecting into the byssal notch, are arranged at wider intervals than proximal inactive ridges near the umbo. This structure is equivalent to Waller's (fig. 3c, p. 211) "row of auricular spines" found in Oxytoma inequivalvis (Sowerby). It is also similar to his second type of pseudoctenolium, called fasciolar, which is formed on the auricular side of the suture and composed of tiny, low and distally pointing projection. The pseudoctenolium of Monotis is different from the fasciolar pseudoctenolium which is exemplified in the shells of Parvamnssiwn sayanum (Dall), because the latter never has more than one active tooth.
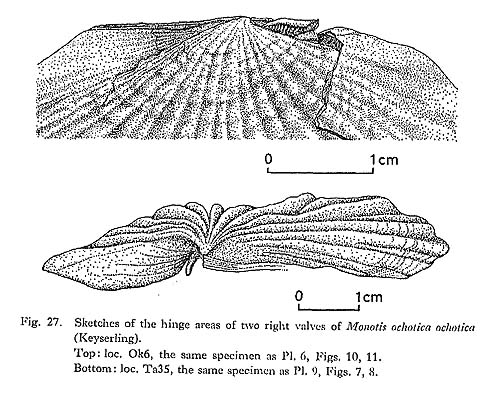
On the lower side of the byssal notch, the anterior disc margin has some coarser growth lines forming short ridges. The ridges number about ten in the specimens of Figs. 21-23 on Pl. 7, and arc arranged from the anterior end of the byssal notch to the middle part of the byssal fasciole. Unlike the byssal notch of O. inequivalvis, this structure has no spines. In spite of the differencc, the ridges of Monotis are probably correlative with Waller's disk-type pseudoctenolium. He commented on its function as follows: "The function of a pseudoctenolium on the disk is unknown, because as yet no one has observed detailed living habits of species having the structure. There does not seem to be any correlation between the occurrence of this type of pseudoctenolium and the strength of byasal attachment. Although the feature occurs only among byssate species, it is more prominent among presumably weakly byssate members of the Aequipecten group, e.g. Cryptopecten, Corymbichlamys, and Aequipecten, than, among strongly byasate members of the Chlamys group (op. cit., pp. 211-212)." In the case of Monotis, however, the notch is so narrow that the pseudoctenolia on the upper and lower sides of the notch possibly have also the function of separating byssal threads and preventing the body from twisting. The anterior part of the auricular ridges, having more conspicuous corrugation than parts nearer the umbo, seems to be the active part for byssal attachment. If Waller's discussions on the derived character state of the ctenolium in the Pectinidae are adopted, the pseudoctenolia in Monotis are suggested to be a primitive character common to other Buchiacean and Pectinacean families and are similar to those of Oxytoma. 4) LigamentAs well as the byssal auricle, the ligament structure is significant for determining the generic and familial position of Monotis (Marwick, 1935; Nakazawa, 1963; Payevskaya, 1985, p. 14). No significant facts amending previous works were obtained in this study, but some comments can be added. Figure 11 on Pl. 6, Fig. 8 on Pl. 9, and text-Fig. 27 show the longitudinally striated ligament area. Following Nakazawa (1963, p. 49), I also question whether "the excavated triangular area which bears about 4 (3 to 6) parallel, longitudinal grooves (Marwick, 1935, p. 258)" is the true ligament pit or not. The specimens used in this study do not have a non-striated and conspicuously excavated pit in both valves. The shallow, striated and triangular excavations observed in both valves are the ligament pits, as was concluded by Marwick and Nakazawa. It is assumed that the Monotis ligament was derived from the typical alivincular ligament by the reduction or modification of the anterior part of the ligament area, as discussed by Nakazawa. The outgroup comparison of the ligament characters with other Mesozoic Buchiacean and Pectinacean genera will provide an important character state for discussing Monotis phylogeny.
2. Intraspecific VariationThe great variability of the outline and ornamentation in each Monotis sample may be partly due to a secondary deformation, but is mainly attributable to wide original intrapopulational variations. Biometrical treatment is useful for the estimation of infraspedfic variation and the analysis of growth pattern. It is applicable to Monotis because of the extremely gregarious occurrence, as long as effects of deformation are taken into consideration. Previous taxonomic studies of Monotis used some statistical methods, e.g. the frequency distributions of univariate characters (including simple ratios) and relative growth analysis (Westermann, 1962; Grant-Mackie, 1978b-d, 1980b-c). Westermann, moreover, adopted the triangular coordinates as frequency polygons, but this approach is not used in the present study. Such bivariate analyses as L-H, B-L and A-L were employed by him, but double logarithmic coordinates were used only for the width to height (H-W) relation. Grant-Mackie employed only ordinary coordinates for such bivariate relations as L-H, A-L, Pt-H, B-(L-A) and W-H. In this study, however, double logarithmic coordinates are adopted for bivariate relative growth, because they are more advantageous in detecting growth-invariant indices (Hayami and Matsukuma, 1970, 1971; Hayami, 1984). 1) Frequency Distributions of Univariate Characters and Simple RatiosFrequency distributions of five selected parameters are shown in Figs. 28-30, though the data of M. mabara arc too poor to be analyzed: M. scutiformis from four areas (Fig. 28); three species and one subspecies from the Shirayamazawa section in the Utatsu area arranged in upward sequence (Fig. 29); several samples of M. ochotica ochotica from four areas (Fig. 30).
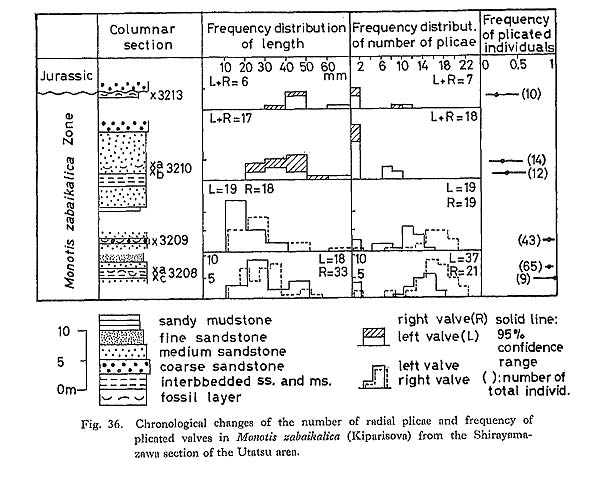
a) Length (L)The frequency distribution of L generally indicates the size distribution which may represent the age composition in a sample. But the scarcity of small valves (less than 10 millimeters) may be attributed to poor preservation and collecting bias. Particularly in M. ochotica size distributions differ from area to area. For example, large valves are predominant in the Okutonc, Takagochi and Nariwa samples, but rather rare in other samples. What factors controlled size distribution are unknown. b) Three Simple Ratios (L/H, (L-A)/A, (L-A)/B)The three simple ratios indicating shell form show wide observed ranges. There is no clear indication of bimodal distribution, though samples of large size are rather scarce. Therefore, these histograms may be mostly interpreted as unimodal, and the hypothesis that each sample is composed of a single phenon is generally acceptable. The H for left valves is a little larger than that for right valves owing to the projected umbo above the hinge line, and the frequency distribution of H/L is slightly different between opposite two valves. The other two ratios do not show differences between opposite valves. M. zabaikalica from the Utatsu area shows that the mode and mean of (L-A)/A and (L-A)/B increase in upward sequence. As the outline of this species becomes more elongate with growth, samples with higher frequency of younger individuals provide smaller values of these two parameters. c) Primary Plicae (Pp)The primary plicae appear at a certain stage of early dissoconch and are rarely bifurcated. Pp is one of the useful growth-invariant characters for the estimation of variations. The histograms of this character in M. scutiformis usually show flat shape and wide range, but it may be partly because of small sample size. In M. ochotica, most of the large samples show similar normal distributions, which may suggest that the sample at each locality consists of only one phenon, Coexistence of a few allied species or a few subspecies in the same species, as previously had been thought by many authors, can never be recognized, so far as the frequency distribution of number of Pp is concerned. The samples of M. ochotica densistriata from the Shirayamazawa section have means of Pp lying between 20.5 and 23, while those of M. ochotica ochotica are between 16.5 and 19.5. Samples of the latter subspecies from other areas have still smaller means between 11.5 and 16.5. At present it cannot be determined whether the difference is due to geographic variation or to chronological change. On the other hand, as shown in Fig. 29, M. zabaikalica has a surprisingly wide dispersion in frequency distribution of Pp. Those histograms are bimodal rather than unimodal, because each sample includes individuals which had no plication from the youngest growth stage. The relative frequency of specimens without radial plicae with 95% confidence range is shown in Fig. 36. The presence or absence of plicae does not necessarily mean polymorphism, because the two conditions are continuous and distinct.
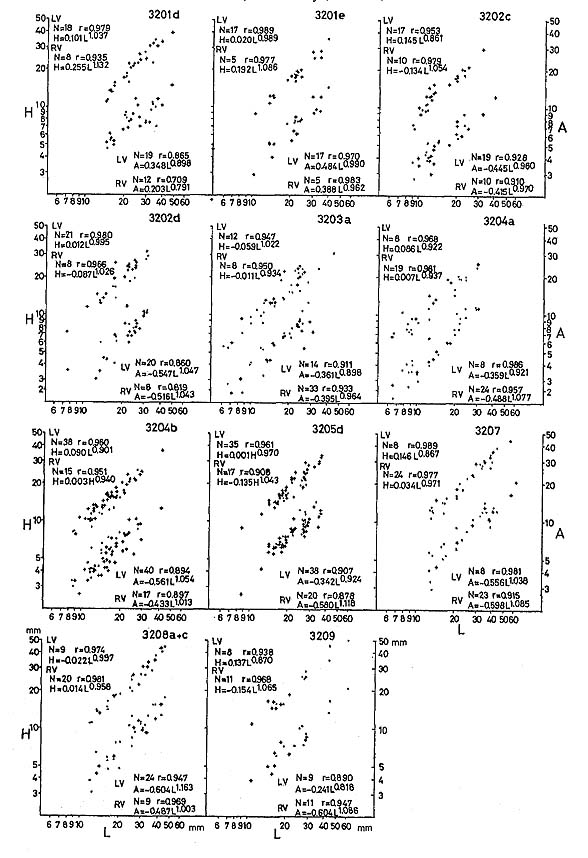
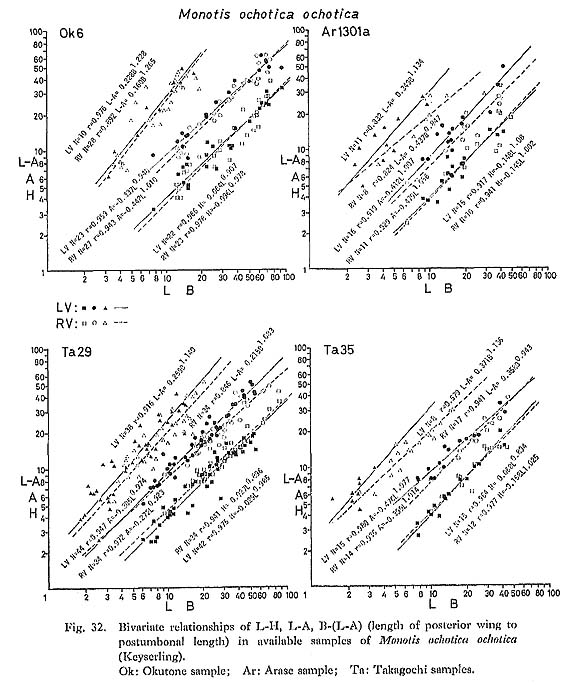
2) Bivariate RelationsThe average relative growth patterns between L and H, and between L and A are shown in Figs. 31 and 32, in addition to the pattern between B and L-A. Logarithmic coordinates and the method of reduced major axis are employed in all samples. It is impossible to recognize a critical point in any case due to wide dispersion. The correlation coefficient between L and H is much higher than between L and A, and L-A and B. This is partly ascribed to the fact that the umbonal part is often deformed.
The dispersion of plotted data is so wide that significant differences of growth ratios between different samples or between two valves can seldom be detected using these three relations. 3) Variation of OrnamentationThe ornamentation of Monotis also exhibits extremely wide variations not only in number, but also in prominence, shape of cross section and insertion pattern of the plicae, which cannot easily be expressed with numerical parameters (Fig. 26). The Takagochi samples are good examples of such variations within M. ochotica ochotica of a late stage (Pl, 9, Figs. 1-8, loc. Ta35; Pl. 9, Figs. 9-22 and Pl. 10, Figs. 1-9, loc. Ta29). In each sample, specimens corresponding to Teller's (1886) five varieties, in addition to the typical form of M. ochotica, and Pseudomonotis sublaevis are included. "Densistriata", "sparsicostata", "eurhachis", "ambigua", "pachypleura", the nominal form "ochotica (s. s.)" and "sublaevis" may be represented by the following illustrated specimens: "ochotica (s. s.)" type individuals: Pl. 9, Fig, 15
Such typological subdivision is difficult to apply to smaller valves of early growth stages. Some right valves with non-plicated umbonal region may be representative of Teller's Ps. sublaevis. M. ochotica poster opiana Westermimn, 1962, which was subsequently described by Kiparisova et al. (1966), is similar to the specimens of Figs. 1-3 on Pl. 10. The problem is how to evaluate these morphological variations. Even if these "taxonomic" names were applied to these morphotypes (or varieties), their boundaries would be quite arbitrary, because the variations must be fundamentally continuous. From the viewpoint of population paleontology, all the specimens of Monotis in each fossil bed seem to represent a single fossil population and cannot be split into taxa. Therefore, it is reasonable to regard the wide morphological variations as intrapopulational. Such varied samples of M. ochotica ochotica were obtained also from the Utatsu (Pl. 5, Figs. 16-20), Arase (Pl. 8, Figs. 1-13), Miyamadani (Pl. 11, Figs. 6-19 and Pl. 12, Figs. 1-16), and Myogatani (Pl. 13, Figs. 1-6) areas.
On the other hand, other samples of M. ochotica ochotica from lower stratigraphic horizons indicate less variable radial plicae with rather regular interspaces (Pl. 5, Figs. 6-15; Pl. 6; Pl. 7, Figs. 1-20; Pl. 8, Figs. 14-24). Besides, M. ochotica densistriata and M. scutiformis show rather regular radial sculpture with wide interspace, though the number of plicae is still variable.
M. zabaikalica, as well as M. ochotica ochotica of late stage, shows a surprisingly wide variation in radial, as well as concentric sculpture (Pl. 13, Figs. 7-20; Pl. 14). For example, the sample from the Utatsu area (loc. Si3208) consists of weakly-plicated and non-plicated valves in the early growth stages. The stage at which the plicae disappear and the pattern of the plicae are variable. At the intersection with a concentric wrinkle at a certain growth stage, most plicae disappear, but some persist till the later stages. Concentric wrinkles also appear variably and irregularly.
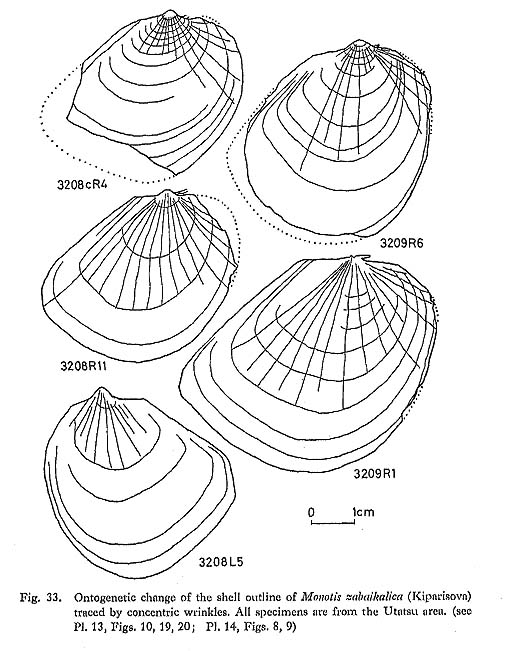
4) Variation of Posterior WingThe shape and ornamentation of the posterior wing are variable, as shown in an example from the Takagochi area (Pl. 10, Figs. 1-3). While one specimen (Fig. 1) shows an obtusely truncated posterior wing, another (Fig. 3) has a truncation angle approaching 90°; Fig. 2 is an intermediate form. The strength of ornamentation on the posterior wing is also quite variable. As exempli fied by the Tsuyama sample, some left valves have a finely plicated wing only obscurely demarcated from the disc (Pl. 7, Figs. 11, 12), while other specimens have a nearly flat and non-plicate wing (Pl. 7, Fig. 8). These two forms cannot be divided by other characters. Therefore, the presence or absence of plications on the posterior wing does not seem to constitute a diagnostic character for higher taxonoinic categories. 3. OntogenyIt is noticed that the outline of the right valve of Monotis at juvenile stages (less than 10 millimeters) is analogous to that of Claraia, Eumorphotis and Oxytoma and the Pectinidae. The outline in the younger stage is nearly subequilateral and subrounded in every species of Monotis treated in this study. In M. ochotica densistriata and M. ochotica ochotica, some right valves lack radial plicae on the umbonal area (Pl. 6, Fig. 9; Pl. 7, Fig. 7; Pl. 9, Fig. 5; Pl. 11, Fig. 3), as described for M. sublaevis (Teller) by Nakazawa (1963, pl. 1, figs, 7-10). These are not thought to be a different species but variants within the same fossil population. Though quantitative analysis has not been carried out, it is found that the ratio of Bl to L decreases with growth, i.e. Bl shows negative allometry with respect to L. On the other hand, the outline becomes more elongate posteriorly with growth in every species. The ontogenetic change of outline is well recognized in M. zabaikalica, because of the presence of prominent concentric wrinkles. Figure 33 shows the ontogeny of the outline worked out by drawing the concentric wrinkles on photographs enlarged two times. Shell margins are sometimes restored as dotted lines by magnifying the earlier wrinkles. Though the outline of juveniles stages (less than 10 millimeters) is aubround or suboval, the shell expands posteriorly and ventrally with growth. As the growth lines are scarcely preserved on the byssal auricles of these specimens, the growth of the byssal auricle cannot be established in detail.
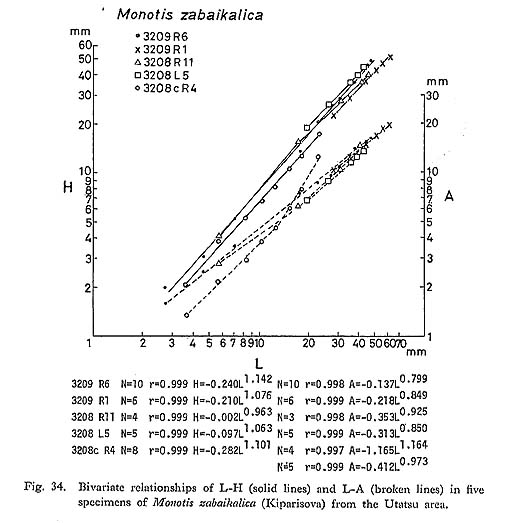
Next, the values of L, H and A in each valve are plotted on double logarithmic coordinates for the analysis of individual relative growth (Fig. 34). The log-log plots of L-H and L-A show nearly linear relations with a certain variance. There is no common critical point recognized in all of these five lines. While the relation of H to L is positive ly allometric, that of A to L is negatively allometric. This means that the ratio of A to L decreases with growth, because of the expansion of the posterior part. The positively allometric relation of H to L may be attributed to the expansion of the ventral part of shell.
In many specimens of M. scutiformis, M. ochotica densistriata and M. ochotica ochotica, the growth lines and concentric wrinkles also indicate posteroventral or posterior expansion with growth. In the later growth stage (over 40 millimeters), M. ochotica frequently has posteroventral sulcation, though the degree and shape are fairly variable (Pl. 6, Figs. 1, 3, 4, 6, 7; Pl. 7, Figs. 24, 25). To sum up, no significant abrupt change in shell characters can be observed during growth. There is no evidence that the mode of life abruptly changed during ontogeny after the shell attained five millimeters in length. |