CHAPTER 7
Geographic Distribution and Faunal Realms
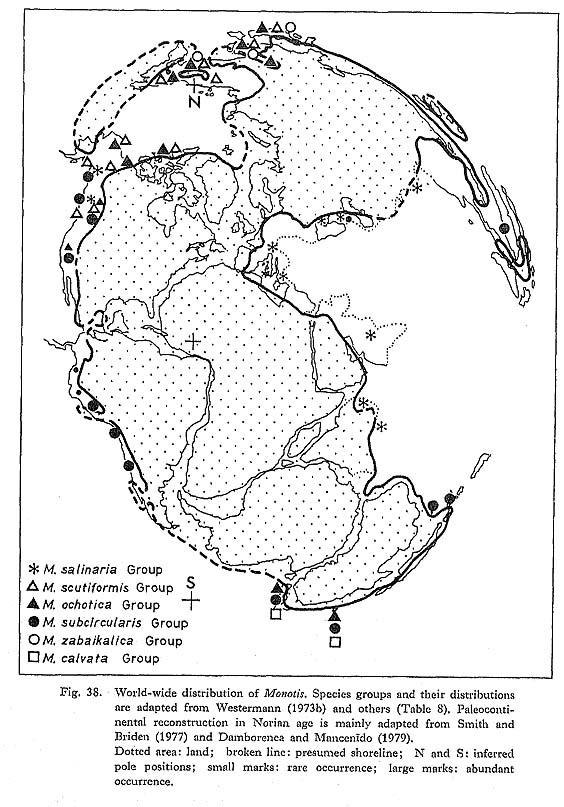
1. Paleobiogeography of Monotis in Japan and East SiberiaThe Monotis fauna everywhere in Japan is usually monospecific, but the constituent species change from horizon to horizon. It does not show any conspicuous provinciality. M. scutiformis has not yet been found from the Outer Zone, and M. mabara does not occur in the Inner Zone of Southwest Japan. This may be due, however, to the incom-pleteness of these Monotis sequences. All the Japanese species and subspecies of Monotis except M. mabara are taxonomically identical with those from east Siberia (especially Verkhoyansk and Kolyma Mountains; Kiparisova et al., 1966). Although the last species, M. mabara, for now, seems to be endemic to Japan, it was probably a daughter species from the main stock which was commonly distributed both in east Siberia and Japan. Whatever horizontal movement of suspect terranes took place, the main part of the Japanese Islands must have been biogeographically connected with east Siberia in Late Triassic times. The faunal con-nection is also well documented by a number of other Carnian-Norian bivalves called the Kochigatani Fauna in Japan, which commonly occur in both regions. In Japan, Monotis occurs in sediments of various grain size from granule-bearing, coarse-grained sandstone to sandy niudstone, in addition to sandy limestone and tuff. The lithology indicates relatively shallow shelf environments. On the other hand, in the Tethyan region, the micritic limestone called the Hallstatt facies is characterized by the occurrence of M. salinaria and is believed to represent a bathyal or pelagic environment. In the Sambosan Belt of the Outer Zone of Southwest Japan, however, the Carnian to Norian faunas are quite different from those of other regions, as pointed out by Tamura (1972, 1981, 1983). They contain obvious Tethyan elements such as Gruenewaldia, Costatoria and Gervillia (Cultriopsis) which show close affinity with species from the St. Cassian Formation in the Alps, though Monotis does not occur there. Recently, Tamura (1981, 1983) discovered megalodontid-bearing limestones with Triadomegalodon sp. cf. tofanae (Hoernes) and Dicerocardium kuwagataforme Tamura, 1983 of Norian or Rhaetian age in eastern Kyushu. He deduced deposition in a lagoon on a volcanic island. Though the distribution of such limestones has been restricted to eastern Kyushu and Shikoku, the writer recently discovered some boulders containing megalodontids at the upper reaches of the Kitaaki River in the western part of Tokyo Prefecture. Therefore, the limestone can be traced to the Kwanto Mountains. Some part of the limestone is somewhat similar to the Hallstatt Limestone, but Monotis has not yet been found. The megalodontid and other bivalve faunas of the Sambosan Belt as well as its sedimentary facies show strong contrast with the Monotis and other Carnian-Norian Kochigatani faunas in the adjacent northern terranes of Southwest Japan. There is a possibility that the Sambosan Belt is a suspect terrane of southern origin and accreted to the northern mainland of Japan. 2. Paleobiogeography of Monotis in the WorldThe worldwide geographical distribution of Monotis is summarized in Fig. 38, assigning various proposed species into six species groups in agreement with Westermann (1973b) and Grant-Mackie (1978a). A list of species groups is shown in Table 8. The M. ochotica, M. subcircularis and M. salinaria Groups are the same as Westermann's, but his M. typica Group is called here the M. scutiformis Group. M. calvata and M. routhieri from the southern hemisphere were included in M. zabaikalica Group by Westermann (1973b), but they are here separated as M. calvata Group (Grant-Mackie, 1976). They were regarded by Grant-Mackie (1978d) as constituting the new subgenus Maorimonotis together with two other species.
As pointed out by Westermann (1973b) and Tozer (1980b), several major Monotis provinces can be recognized; for example, the Western to Northeastern Pacific province is characterized by the M. ochotica Group, the Eastern Pacific province by M. subcir-cularis, the Tethyan province by M. salinaria, the Northwestern to Northeastern Pacific province by M. scutiformis, the Northwestern Pacific province by M. zabaikalica, and the Southwestern Pacific province by M. calvata. Among others, M. scutiformis, M. ochotica and M. zabaikalica, as described in this paper, seem to constitute an evolutionary lineage. Recently, Tozer (1982) discussed the distribution of Triassic marine, invertebrate faunas in North America considering plate and terrane movements. Current develop-ment of plate tectonic theory has presented an interpretation in which the greater part of the North American Cordillcran margin is actually a mosaic of terranes which "seem to have been swept from far reaches of Pacific Ocean before collision and accretion into the Cordilleran margin mostly in Mcsozoic to early Cenozoic time (Coney et al., 1980, p. 329)" (Tozer, 1982, p. 1080). He adopted the term "suspect" for the exotic terrane whose origin is uncertain, and the term "plate-bound" for the terrane that was de-posited on a continental plate in situ. The North American Cordilleran faunas were classified into three types, namely the low, mid and high paleolatitude faunas (LPL, MPL, HPL) regardless of being plate-bound or suspect, Monotis occurs both in plate-bound sequences and in suspect terranes. Tozer thought that M. salinaria in the Hallstatt Limestone facies was an indicator of LPL, M. subcircularis of MPL or LPL and Eomonotis (M. scutiformis Group in this paper) and M. ochotica of HPL or MPL, con-sidering the faunal composition, sedimentary facies and geographical distribution of faunas in Westermann's scheme. During the preparation of Fig. 38, the need arose to explain various disjunct distri-butions of Monotis species; for example, M. subcircularis in Borneo (Tamura and Hon, 1977), M. salinaria in Mexico and east Siberia, etc. These distributions could be ex-plained if the collision and accretion of suspect terranes were accepted, but this is pos-sibly an untestable hypothesis. Recently, Grant-Mackie (1978a-c, 1980b, c) described a number of Monotis species (Table 8) from New Zealand, Though detailed comparison remains to be done, I pre sume that some species of Eomonotis and Entomonotis proposed by him are synonymous with, or closely related to, the east Asian species. Because the Maorian monotids were too finely split, the true diversity may not be so high. Wider assessment of the intra-specific variation appears to be needed. Other Maorian bivalve faunas of Carnian to Rhaetian age indicate considerable differences from the Northeastern Pacific and Tethyan regions, as was summarized by Kobayashi and Tamura (1983a, b). |