CHAPTER 4
Mode of Occurrence and Ecology of Monotis
|
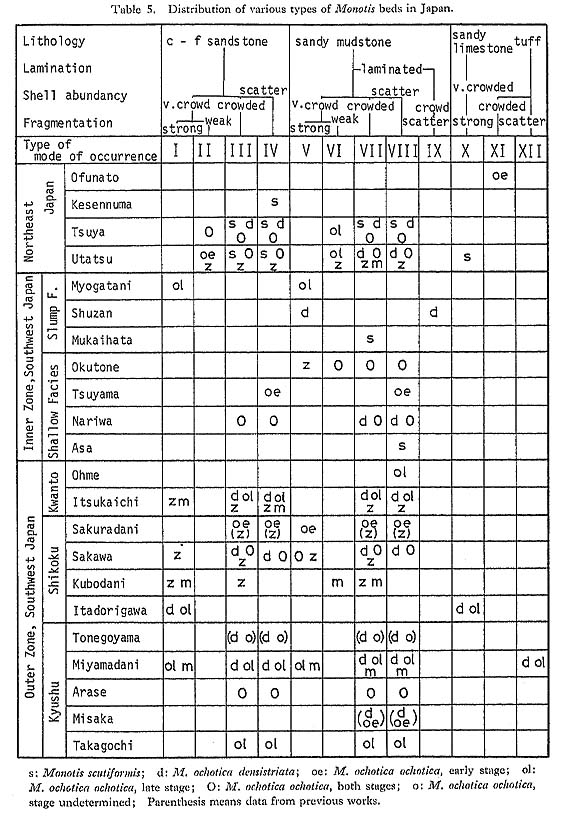
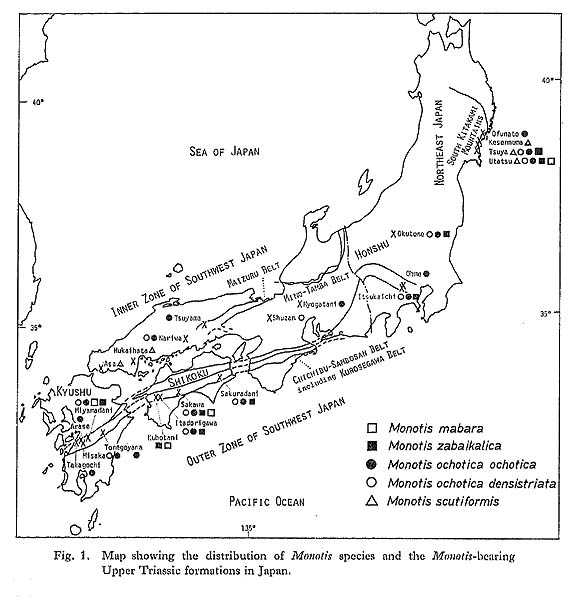
Although some authors paid attention to the stratigraphic occurrence of Monotis, little has been said about the mode of fossil occurrence in each fossil bed. The mode of fossil occurrence may furnish valuable clues to the paleoecology of Monotis, where and how it lived. Before going into further disucussion, results of observations in the field and laboratory arc mentioned, with reference to some previous observations on this problem. 1. Criteria for Evaluation of Mode of Occurrence of MonotisIn order to recognize the mode of occurrence of Monotis shells, the fossil beds at various localities in Japan were sedimentologically and taphonomically examined. Several samples were collected as slabs or blocks for laboratory observation. The sedimentary structure was studied by examining polished sections cut vertically and horizontally to the bedding plane. Sedimentary basin analysis over a wider area may be important, but it was often prevented by the complicated geologic structure, poor exposures and the scarceness of good key beds. As shown in Table 4, twelve types, I to XII, were distinguished by such criteria as rock type, lamination, shell abundance and shell fragmentation, because these seem to be related to the mechanism of the formation of fossil beds. Table 5 shows how Monotis species are distributed in each area in relation to the twelve modes of shell occurrence. The mode of occurrence of Monotis shells is described in the following sections with regard to these criteria.
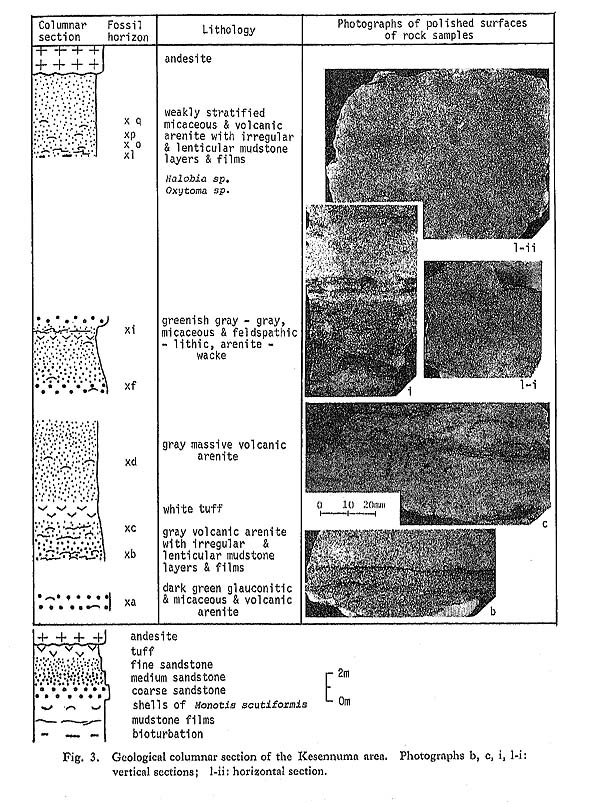
1) Rock TypeIn Japan, Monotis occurs in sediments of various grain, size from granule-bearing, coarse-grained sandstone to sandy mudstone, in addition to sandy limestone and tuff. a) Sandstone Sandstone, yielding Monotis shells, is either of wacke or arenite type, lithic to feldspathic, poorly sorted, and frequently with a large amount of mica, volcanic rock fragment, mud and mud patch. It is, in general, sedimentologically immature. Crowded Monotis beds are frequently calcareous and rarely glauconitic (e. g. loc. Ke-a, Kescnnuma area; Fig. 3). The mode of shell occurrence in sandstone is classified into four types, I to IV, by the combination of two other criteria (Table 4; Pl, 1, Figs. 1-4).
b) Sandy Mudstone Mudstone is also a common rock type for Monotis beds in Japan and probably also in east Siberia (Kiparisova et al., 1966; Gromov and Tuchkov, 1971). It is mostly sandy and silty, frequently very micaceous, poorly stratified, poorly laminated and slightly to conspicuously bioturbated. It is rarely glauconitic (e.g. Iocs. Sr4103c, 3213, Utatsu area; loc. Ku10, Kubodani area). Abundance of bioturbation and absence of lamination suggest deposition on a shallow-sea bottom where some benthos lived and disturbed the substratum. This facies includes types V to IX (Pl. 1, Figs. 5-8; Pl. 2, Figs. 1-6). Type IX shows exceptionally conspicuous lamination which consists of thin alternations of sandy and muddy laminae (Pl. 2, Figs. 7-9).
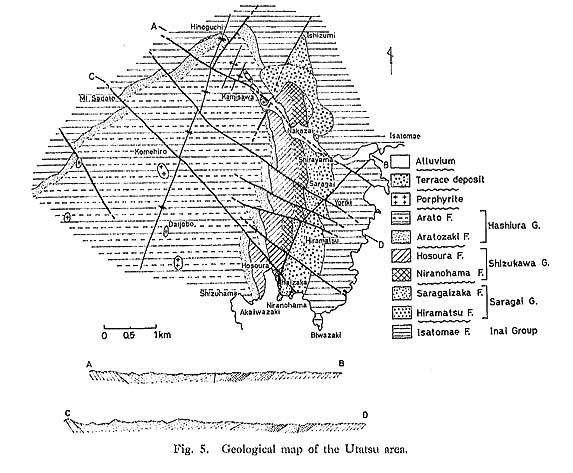
c) Sandy Limestone In the Utatsu and Itadorigawa areas, sandy limestone forms very crowded shell, beds which contain numerous fragmentary Monotis shells, skeletons of crinoid and foraminifers, detrital sand grains and volcanic rock fragments (Pl. 3). The Utatsu sample of sandy limestone, particularly, contains a large amount of mud. The Itadorigawa sample contains some lenses of light greenish tuff.
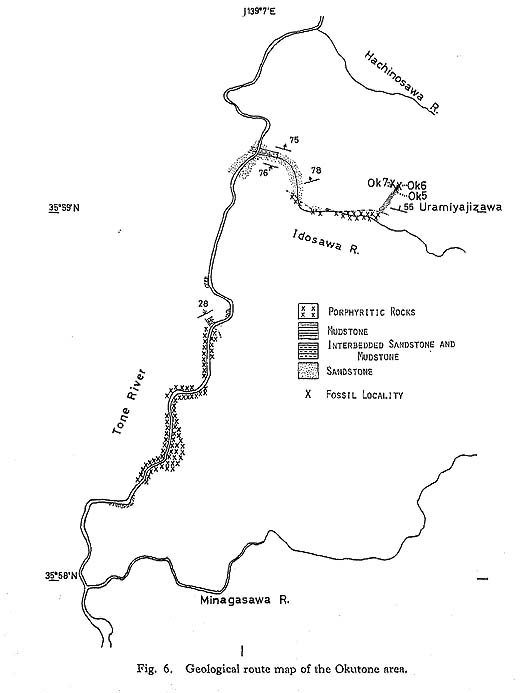
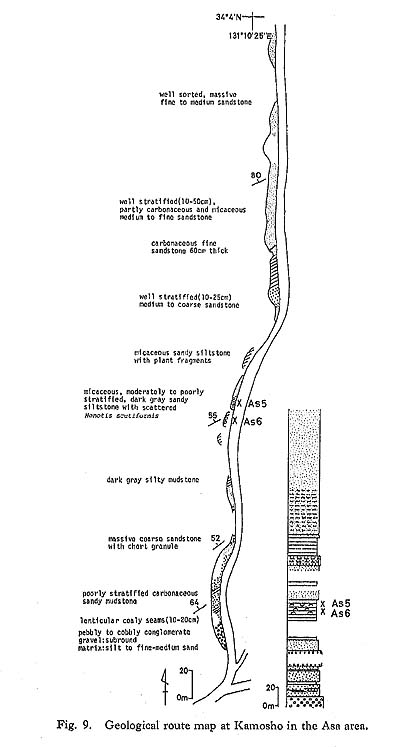
d) Tuff The rarest rock type in the Monotis beds in Japan is tuff, which occurs as thin intercalations in the shallow-sea sediments. Such tuff layers appear to have been transported by currents from some primary depositional site, as exemplified by the Monotis bed in the Miyamadani area (Fig. 21).
On the other hand, the Ofunato sample shows a unique mode of occurrence, because the tuff bearing crowded Monotis shells is contained as pebbles in volcanic breccia. 2) LaminationLamination is generally undeveloped in the Monotis beds though horizontally arranged shells themselves constitute laminae in the beds of types I, II, V and VI. It is sure that some benthic organisms disturbed the original laminations. This criterion discriminates only type IX from others. Well developed laminations are observed only in samples of the Shuzan area (Pl. 2, Figs. 7-9). 3) Shell AbundanceThree grades, that is, very crowded, crowded and scattered, are applied though only qualitatively, to express shell abundance. The highest grade, "very crowded", indicates a very condensed shell bed in which numerous shells and their fragments are laid upon others or side by side. The thickness varies from ten centimeters to several meters. The thickest shell beds of this type are observed at Tsuya (loc. Ch1486; Pl. 1, Figs. 7, 8) and Okutone (loc. Ok5; Pl. 1, Fig. 5) areas. The second grade, "crowded", indicates that several to many shells are frequently laid upon others or side by side, but are not so condensed as the first grade. Patches containing several shells are frequently scattered on a bedding plane. Monotis beds of this grade are the most frequent. The third grade, "scattered", represents occurrences where a solitary or a few shells are scattered over a bedding plane. Sometimes patches containing several shells are scattered. This grade is also common. In every grade of shell abundance the Monotis shells seem to have been deposited and concentrated by water currents, since in situ occurrences have not been observed at any locality in Japan. This gregarious and exclusive shell occurrence may be related to the opportunistic strategy of Monotis (Westermann, 1973b; Hallam, 1981), though there is no reliable evidence. 4) FragmentationSince Monotis shells are very thin and apt to be broken during transportation, every shell bed contains numerous fragments. Two grades of fragmentation are distinguished in very crowded layers, namely strongly fragmented layers represented by types I and V, and weakly fragmented ones by types II and VI. Fragmentation is dependent upon the rock type. For example, the frequency of fragments is generally higher in sandstone than in sandy mudstone. Type II, characterized by sandstone and very crowded shells, however, includes a relatively small number of fragments (Pl. 1, Fig. 1). 5) BioturbationThe sandstones and mudstones of Monotis-bearing formations commonly show bioturbated structures such as heterogeneous mottles, disturbed laminae, pelletal grains, and horizontal muddy excrements. The first two structures are very frequently observed in poorly sorted, muddy fine-grained sandstone and sandy mudstone of types III, IV, VII and VIII in many areas (Pl. 1, Figs. 3, 4; Pl. 2, Figs. 2-6). Massive sandy mudstone bearing scattered Monotis shells in every area shows some bioturbation. Very crowded shell beds, however, rarely exhibit bioturbation, regardless of the lithology. The horizontal muddy excrement is classified into two types. The first type is represented by the muddy excrement on a bedding plane of poorly sorted, medium-grained sandstone in the Kesennuma (Fig. 3 l-i, ii) and Utatsu (Fig. 22) areas. It is composed of bent chains of a few to several globular mounds or rounded squares, 2 to 4 millimeters in diameter, or solitary globules, which are gray mud and different from the host rock in lithology. There are no mud films or layers of the same lithology, above and below the bedding plane. In the Utatsu sample the excrement is found to be associated with several Monotis shells. Whether the solitary mounds are primary or represent chains broken by current or biogenic disturbances, cannot be determined at present. Judging from the restricted occurrence on a plane and the lithology of the host rock, they do not appear to be burrows produced by an infaunal animal but may be excrement left by epifauna or infauna at periodic intervals on the sea floor.
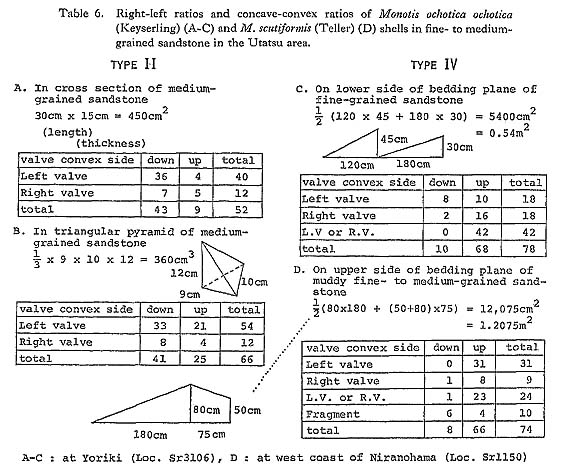
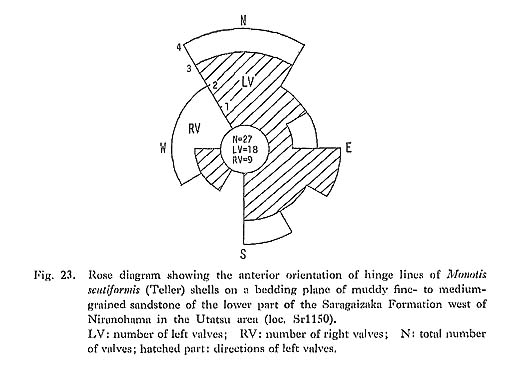
The second type of horizontal muddy excrement similar to the first, was obtained from gray sandy mudstone which bears a few scattered Monotis shells in the Miyamadani area (Pl. 2, Figs. 5, 6). Though similarly composed of bent chains of globular mounds or rounded squares, it differs from the first type in the size and the lithologies of mounds and host rock. The size is four to six millimeters in width or diameter. The lithology of the excrement is black clay, while that of the host rock is gray sandy silt with pelletal clayey grains. Moreover, whereas the first type occurs only on a particular bedding plane with many solitary mounds, the second constitutes a layer more than several centimeters thick with no solitary mounds. Laminae in the host rock are obscurely curved around the mounds. It is inferred that this excrement was left also by epifauna or infauna. The two types of excrement are very similar to the trace fossils named Toyonereites eitaroi Katto, 1962, which was in the sandy shale 20 meters above the bed with Monotis scutiformis in the Asa area. Taking the size and the lithology into account, Toyonereites eitaroi is probably an equivalent of the second type from the Miyamadani area. At present there is no reliable evidence that bioturbation produced by benthic animals in soft sediments was related to the removal, transportation and accumulation of Monotis shells. 6) Mica ContentMonotis-bearing formations, both sandstone and mudstone, generally contain rather a large amount of mica flakes indicative of sedimentological immaturity. The abundance of mica appears to suggest a shallow-aea environment where detrital grains were in abundance. 7) ConvexityBowl-shaped single valves of organisms such as bivalves and brachiopods commonly occur preferentially in convex/concave positions, depending upon the fluid regime and sedimentary process in which they are deposited (Müller, 1979). Stronger convexity of left valve of Monotis is one of the reasons why differential sorting took place. Except type II, convex-up position generally dominates over convex-down position, though very crowded beds such as types I, V and VI are difficult to analyze due to fragmentation and clumping of shells. Sandstone with scattered shells, type IV, has shells in the convex-up position more frequently than type III with crowded shells (Table 6). Figure 23 shows the anterior orientation of hinge lines of M, scutiformis on a bedding plane of the same sandstone as in Table 6D. Only the obscure orientation in NNW-SSE can be observed due to the subrounded shell outline. In type II, nevertheless, the convexdown position of left valve is predominant (Pl. 1, Fig. 1). The shell bed, about 25 centimeters thick, is intercalated in massive to thickly bedded, medium-grained sandstone with scattered shells.
8) Ratio of Left and Right ValvesAs it is difficult to extract many valves of Monotis from hard rocks, the relative frequency of left and right valves cannot be easily determined. In general, the right valve of Monotis seems to be weaker than the left which has stronger convexity, even if the two valves have the same shell thickness and ornament. The right valve is apt to be broken selectively prior to burial. In fact, the left valves are almost always dominant in sandstone, regardless of the shell abundance, while a ratio of about 1: 1 is generally maintained in mudstone. 9) DisarticulationProbably due to the weak ligament and the absence of teeth, Monotis valves seem to be easy to disarticulate. In fact, they are almost always disarticulated even in mudstone. Only at loc. Sr 1050 in the Utatsu area, were some conjoined valves of M. scutiformis occasionally found. 10) Faunal Composition and Associated FaunaOne of the striking features of Monotis beds is that other associated fossils are extraordinarily rare. Monotis shells represent more than 99% of total individuals in a shell bed, commonly as a monospecific fauna. Among the rare associates, the most frequent fossils re brachiopods, followed by such epifaunal bivalves as Oxytoma and Halobia, the nfaunal bivalve Palaeoneih and epifaunal crinoids. The sandy limestone in the Itadorigawa area contains a small amount of foraminifers (Pl. 3, Figs. 2, 7, 8). 2. Sedimentary Environment of Monotis Beds in JapanThe lithology and the mode of fossil occurrence described above indicate that Monotis beds in Japan are neither offshore nor deep-sea sediments, but surely of near-shore or shallow-sea origin. This is in considerable contrast with Daonella and Halobia, which always occur in finer and probably more offshore sediments. Hallam (1981) pointed out the apparent paradox between the opportunistic strategy of Monotis inferred from its gregarious and exclusive occurrence and the stable offshore sedimentary environment inferred from the lithofacies. This is probably true in the case of M. salinaria, which is contained in micritic limestone or calcareous mudstones of the Tethyan province called the Hallstatt facies. Japanese Monotis beds are quite different from the Hallstatt facies in the Alps in the dominance of clastic sandy sediments which indicates higher sedimentation rates. Sandy limestones in the Utatsu and Kubodani areas may have been formed in a shallow-sea environment where a great number of fragmentary Monotis shells were deposited with detrital grains, fragmentary skeletons of crinoids and foraminiferal tests. In the Utatsu area the sandy limestone is overlain by cross-bedded sandstone with a rather sharp lower boundary. Though there is no evidence of tempestites (Aigner, 1982), the Monotis shells seem to have been rapidly deposited by rather strong currents or storms. The sandstone of type II may represent a short-event deposit because Monotis shells occur dominantly in the convex-down position. The sandstone of types III and IV and sandy mudstone of types VII and VIII are so bioturbated that some benthic animals must have lived in and disturbed soft substrata. Monotis shells are generally 50 to a few hundred microns in thickness, but are still frequently preserved even in sandy mudstone. The Monotis beds in sandy mudstones do not show bituminous lithology anywhere in Japan. Therefore, the sedimentary environment of the Japanese Monotis beds was never an oxygen-deficient, stagnant bottom. Next, the "slump" facies in the Shuzan area will be mentioned. In the Shuzan area, the Monotis bed, 3 meters thick and composed of alternating thin sandy and muddy laminae several millimeters thick, may have been deposited in an offshore basin where slumping took place, if the relation between the Monotis bed and the slump breccia is conformable. The Monotis-bearing part, however, contains no major-acale slump structure. This may represent the deepest facies among the Japanese Moiiotis beds. The total thickness of the Shirayamazawa section in the Utatsu area is estimated to be 150 meters. However, the Monotis section on the west side of the Okhotsk Mountains (Gromov and Tuchkov, 1971) attains a thickness of 450-500 meters, despite the M. zsbaikalica Zone not being represented. The most important section in New Zealand is the Kiritehere coastal section which is approximately 75 meters thick without the part of jumbled small packets (Grant-Mackie, 1980d, 1985). The Japanese Moiiotis beds, as well as the Okhotsk and Kiritehere sections, are mainly composed of sandstone and mudstone, and the thickness seems much larger than that of limestone facies such as the Hallatatt Limestone in the Alps and the Pine River Bridge section in British Columbia. For example, the thickness of the Pine River Bridge Section in British Columbia is only about 23 meters in the thickest part (Westcrmann, 1962). 3. Paleoecology of MonotisTable 7 shows the modes of life of seven Mesozoic thin-shelled bivalve genera as inferred by several authors. These are mainly based upon circumstantial evidence such as mode of shell occurrence and functional shell morphology, because there are neither extant relatives nor fossil occurrences directly showing the mode of life. At present the paleoecology of Monotis must be inferred from various lines of indirect evidence.
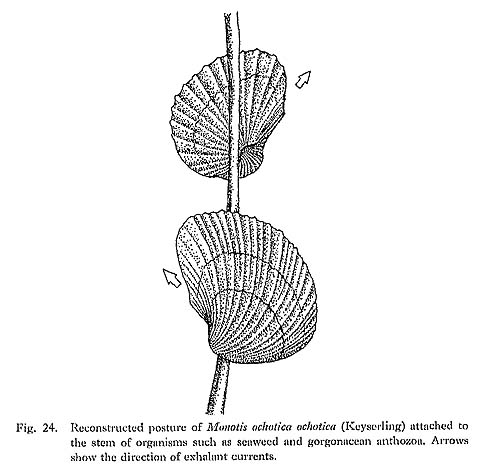
Among various works, excellent discussions were given by Ichikawa (1958), Jeffcries and Minton (1965), Hayami (1969), Westermann (1973b) and Grüber (1976) on the mode of occurrence and ecology of Monotis. The two most likely inferences were for a pseudoplanktonic and benthic mode of life. Since Schmidt (1935) inferred a pseudoplanktonic mode of life for Daonella, Halobia and Monotis, such a inode of life, which has not been recognized in extant bivalves, was accepted by others (Ichikawa, 1958; Weatermann, 1962, 1973a, b; Hayami, 1969, etc.). Other authors thought them benthic (Krumbeck, 1924; Stanley, 1972; Grant-Mackie, 1980a; Hallam, 1981). Recently, Grant-Mackie (1980a) made a more daring inference for Monotis ecology. According to him Monotis may have been epifaunal and attached itself to sessile seaweed. M. (Eomonotis) may have adhered to the distal portion of the seaweed laminae and M. (Entomonotis) more to the proximal portion. On the other hand, morphologically aberrant M. (Maorimonotis) may have been a true bottom dweller and endobyssate. Periodic storms may have swept the weeds with attached epifauna to other places, allowing Eomonotis and Entomonotis to be distributed widely as accidental pseudoplanktonic individuals. Some of them may have succeeded in reproducing and establishing new populations. There is no positive evidence for such a suggestion, though it is attractive because it concerns slightly different niches for the three subgenera. As Japanese Monotis species, which are mostly the same as east Siberian species, have inequivalve and inequilateral shells and a distinct byssal auricle on the right valve, they cannot be free swimmers such as Pecten and Lima or burrowcrs. They could not have moved along a straight path in water. The presence of the anterior auricle and a narrow, but deep, byssal notch is noteworthy. Finely transverse ridges can be observed on both sides of the notch. These ridges, called pseudoctenolia, seem to have functioned as ctenolium, as described in p. 56. Moreover, Monotis has extraordinarily thin shells and shows a negatively allometric relation of shell thickness to length (or shell size). Therefore, the byssal attachment may indicate an epizoic mode of life. If so, to what organism or object did Monotis attach itself? In spite of many observations of Monotis beds, neither the organisms nor the objects can be found. No remains and traces of attached epifauna on the shells of Monotis are known. Taking sedimentary fades into account, some sessile organism seems to be a plausible object to which Monotis may have attached, so far as Japanese Monotis is concerned, and as inferred by Grant-Mackie (1980a). If pseudoplanktonic, fossil occurrence would be independent of sedimentary facies. Monotis should occur both in shallow-and deepsea facies, regardless of bottom environment. Japanese Monotis, however, is restricted to shallow-sea sediments which seem to indicate a well-oxygenated environment. At least there are no undoubted deep-sea sediments with Monotis in Japan. The auricle is too minute to have several such functions as supporting attached shells, as in pectinids, and postulated sheltering in Pteria as recognized by Stanley (1970, fig. 12). Assuming byssal attachment to the stem of benthic seaweed or gorgonacean anthozoa, the living position of Monotis is reconstructed in Fig. 24. As the bysaal auricle is situated close to the umbo and the valves are strongly inequivalve, the sagittal plane may not have been vertical. Then the projection of the convex left valve possibly accomplished a supporting function. A more strongly convex left valve may have been directed obliquely downward with the posterior margin of the posterior wing obliquely upward. But no shell torsion for the supporting function can be observed in the umbonal part of any specimen. The exhalant water may have been expelled from the posterior margin of the wing.
The ecology of M. salinaria and M. subcircularis must be considerably different from that of the Japanese species discussed here, because they have subequivalve shells and generally occur in more offshore sediments. These species were probably offshore and may have occupied the niche which had been occupied by Daonella and Halobia in. earlier times. |