CHAPTER 3
Stratigraphic Distribution of Monotis
|
As Monotis is a very useful fossil index of Norian age, its Stratigraphic distribution has been studied by a large number of stratigraphers and paleontologists, namely Shimizu and Mabuti (1932), Kobayashi and Ichikawa (1949), Ichikawa (1954b), Nakazawa (1963, 1964a, b) and Tamura (1965) in Japan, Westermann (1962, 1966, 1973a, b), Westermann and Verma (1967) in northeastern, British Columbia, Kiparisova et al. (1966) and Gromov and Tuchkov (1971) in east Siberia, and Grant-Mackie (1976, 1978a, 1980d, 1985) and Campbell and Grant-Mackie (1984) in New Zealand and New Caledonia. These works were summarized by Westermann (1973b, table I), Tozer (1980b) and Grant-Mackie (1985) (Table 1).
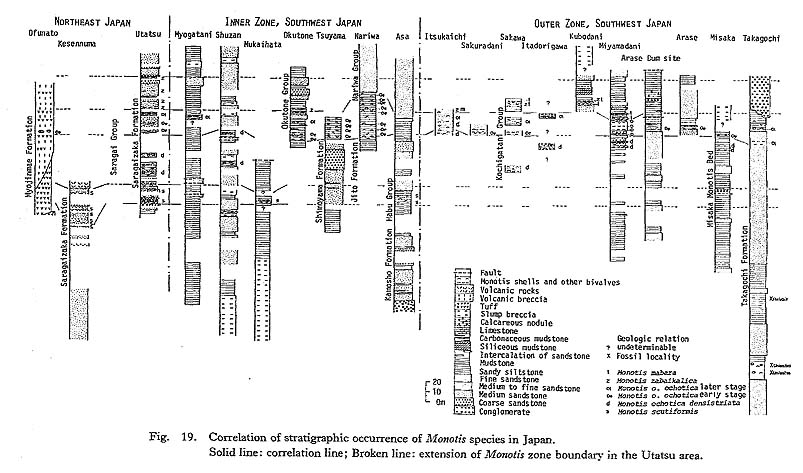
With regard to the precise age of the first and last occurrences of Monotis, little new information has been obtained in this study. The exclusive occurrence of Monotis shells and the scarcity of other index fossils make it difficult to check the contemporaneity of Monotis zones. In particular, though Japanese Monotis beds very rarely yield a few ammonites such as Arcestes, Rhacophyllites and Placites, they are not identifiable at the species level, because of the poor preservation. On the other hand, conodonts, which occur abundantly from the Triassic cherty sediments, are seldom found in shallow-sea sediments such as the Japanese Monotis beds. So it is still difficult to correlate the Monotis zones with conodont zones. The Okuku Limestone and the Mount Mason Limestone of the Torlesse Super Group in the South Island of New Zealand contain some Norian conodonts, such as Paragondorella navicula navicula (Huckriede), Paragondorella steinbergensis Mosher, Hindeodella suevica (Tatge) together with Monotis richmondiana (Jenkins and Jenkins, 1971). Although correlation with conodont zones may be possible in limestone facies, the coexistence of Monotis and conodonts has not been found in Japan. Therefore, I provisionally adopt the Stratigraphic range of Monotis discussed, by Westermann (1973b) and Tozer (1980b). But there is a problem as to what kind of chronological scale should be applied for the interval between the Upper Triassic Himavatites columbianuis zone and the basal Jurassic. This problem was discussed by Tozer (1979, 1980a, b), Wiedmann et al. (1979), Harland et al. (1982) and Isozaki and Matsuda (1983). It has, however, not been settled yet. Tozer (1967, 1979, 1980b) concluded that Monotis occurs only in. two ammonite zones, i. e. M. scutiformis in the Himavatites columbianus Zone and M. ochotica, M. salinaria, and M. subcircularis in the Gnomohalorites cordilleranus Zone in British Columbia. According to Harland et al.'s age determination, the absolute time range of Monotis can be estimated to be only about a few to several million years. If the Rhaetian is eliminated as an independent stage, which is supported by Tozer (1979) and Hallam (1981), Monotis appeared at the late Middle Norian and became extinct during the Upper Norian. Following the alternative recommended by the IUGS Subcommission on Triassic Stratigraphy (Richter-Bernburg in Wiedmann et al., 1979), it ranges from the Late Norian to the Early Rhaetian in age. Which is better, is beyond the purpose here. Stratigraphically documented serial samples of Monotis, which are indispensable for evolutionary study, have been looked for throughout Japan. Figure 19 shows the outline and correlation of successions in Japan. Successional samples were obtained in only a few areas, as discussed below.
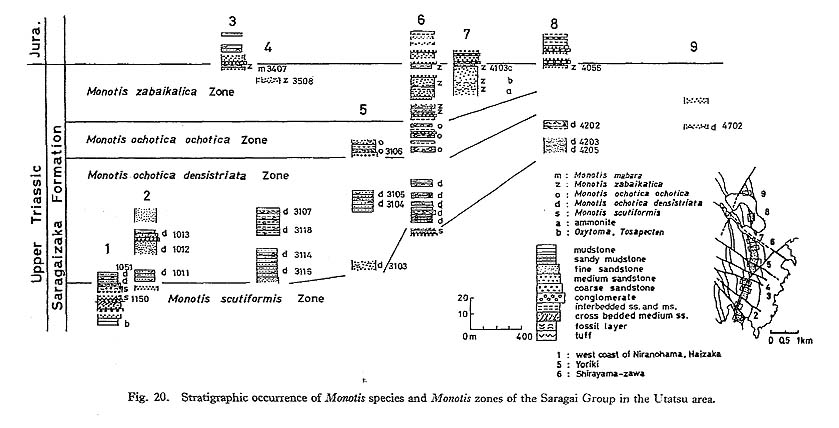
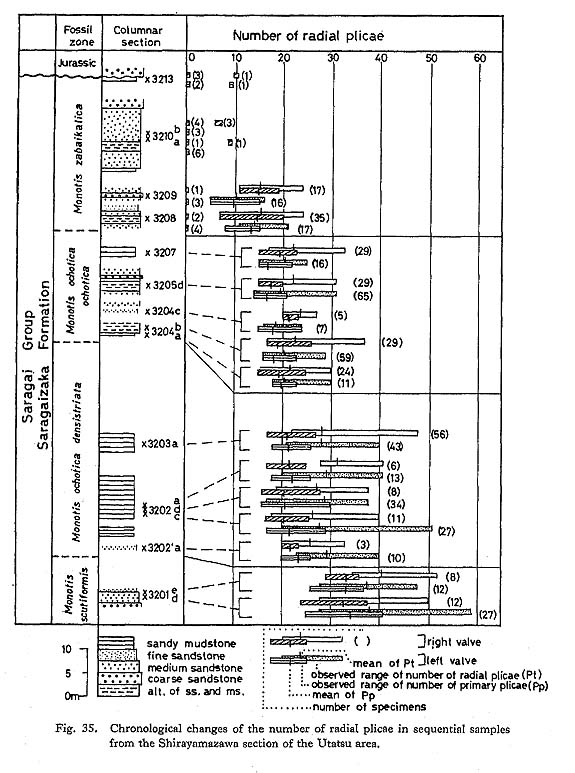
1) Utatsu AreaIn the Utatsu area, a nearly complcte succession of Monotis is observable. Among others, the section along a small valley, called Shirayamazawa, is most useful (Fig. 20). The standard Monotis zonation adopted in this paper is primarily based on this section. Samples from 23 horizons were collected for population studies. M. scutiformis (Teller), M. ochotica densistriata (Teller), M. ochotica ochotica (Keyserling), M. zabaikalica (Kiparisova) and M. mabara (Kobayashi and Ichikawa) [= Entomonotis tenuicostata Kobayashi and Ichikawa, 1949; non Monotis tenuicostata Kittl, 1912; see p. 92] in upward sequence were recognized, taking intrapopulational variation into account. The first appearance of each species or subspecies, except for M. mabara defines a fossil zone traceable to an other section along the eastern limb of the Shizukawa Syncline. Generally, the zone boundaries seem to correspond well with the lithological boundaries, The two chronological subspecies of M. ochotica are Stratigraphically separated but similar in morphology except for shell ornamentation (Fig. 35). Therefore, they are arbitrarily divided at a certain horizon. Similarly, M. scutiformis, M. ochotica densistriata, M. ochotica ochotica, and M. zabaikalica seem to constitute individual lineage zones, although the uppermost part of the M. ochotica ochotica Zone yields a small number of shells resembling M. zabaikalica (loc. Sr3207). As all the specimens from the next fossil bed at loc. Sr3208 are assigned to true M. zabaikalica, it is inferred that the descendant morphology of M. zabaikalica is anticipated in the population of M. ochotica ochotica. The last zone is dominated by M. zabaikalica, but M. mabara is very rarely found in association with M. zabaikalica at loc. Sr3407.
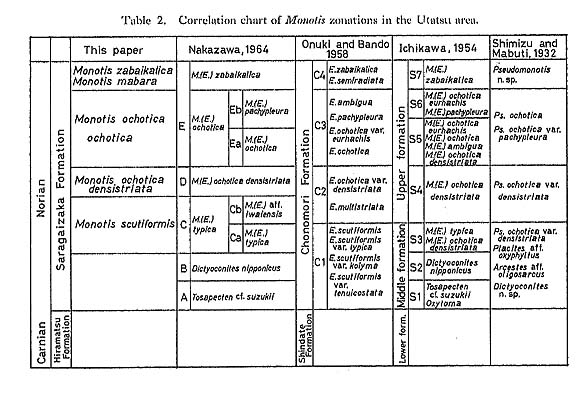
The zonal scheme confirmed here is shown in Table 2. The present quadripartite division may seem similar to that of Onuki and Bando (1958; C1 to C4 zones) and also to that of Nakazawa (1964a, b; C to E zones plus the M. zabaikalica Zone). The constituent species characterizing each zone are, however, considerably different. In their works each zone was characterized by the co-existence of one dominant species and a few subordinate species (or subspecies). For example, Ichikawa's (1954b, 1958) zoning seems to have been based on the assemblages of several morphological species or subspecies. He recognized seven Monotis zones by their relative frequencies. As pointed out by Hayami (1969, 1975), it is biologically unlikely that so many related species and subspecies would occur together, even if dead shells were transported.
2) Miyamadani AreaFigure 21 indicates the section in the Miyamadani area. The continuous exposure about 32 meters thick yields abundant Monotis shells. It provides Monotis samples from 23 horizons, though the sample sizes are highly variable because of different shell densities (Pls. 11, 12). The lower six horizons, a to f, contain shells with finer primary and numerous higher-order plicae, which are referable to M. ochotica densistriata. These samples are slightly different from those from the Utatsu area in the smaller number of primary plicae. The middle eight horizons, i to p, contain numerous specimens of M. ochotica ochotica, which are different from the Utatsu specimens in having fewer and thicker primary plicae with a smaller number of higher-order ones. The upper portion is characterized by M. mabara. The finely-plicate individuals of M. ochotica ochotica at horizon p resemble ordinary individuals of M. mabara at horizon q. But the two samples are distinguishable from each other as a whole.
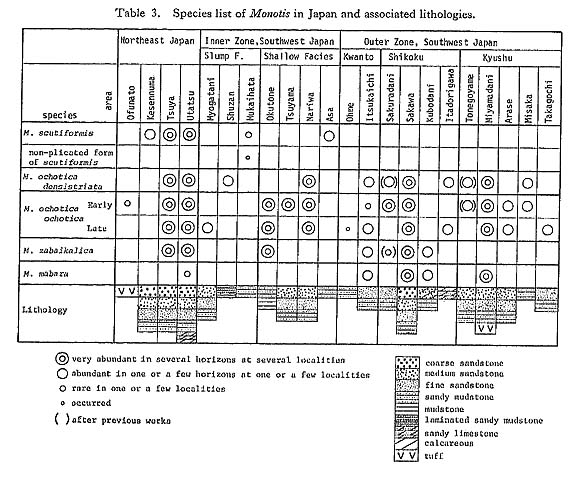
Tamura (1965) recognized four fossil zones, that is, M. sabaikalica, M. pachyplcura, M. multistriata and M. tenuicostata Zones in upward sequence above the M. ochotica (s. s.) Zone. As a result of my survey, no specimens exactly identifiable with M. sabaikalica were found. The portion of the section above the M. ochotica ochotica Zone could not be divided into zones, but should be assigned to a single zone characterized by M. mabara, though the specimens are poorly preserved and often fragmentary. 3) Sakawa AreaIn the Sakawa area there are many Monotis localities, as previously reported by Kobayashi and Ichikawa (1949, 1950). The sporadic exposures, poor state of preservation and extraordinarily complicated geologic structure, however, make it difficult to detect an undoubted Monotis sequence. M. ochotica densistriata and M. ochotica ochotica occur separately at some localities, while M. zabaikalica and M. mabara occur together at others. As pointed out by Kobayashi and Ichikawa (1949, 1950), Monotis faunas can be subdivided into two subfaunas, namely M. ochotica-M. densistriata and M. tenuicostata (=M. mabara in this study)-M, zabaikalica subfaunas. They indicated the two subfaunas do not occur together except at one locality. This mutually exclusive pattern of occurrence is also confirmed in this study. Various partial and incomplete Monotis sequences in Japan can be compared with the standard Shirayamazawa section in the Utatsu area (Fig. 19), although the absence of any good key bed and index fossils such as ammonites and conodonts, makes accurate correlation difficult. The Monotis-bestring part does not exceed 150 meters in thickness in any area. The Monotis succession confirmed in the Utatsu area seems to be compatible with other areas in Japan. In other words, the following succession of stratigraphic occurrence, though often incomplete, is invariably observed throughout Japan: Monotis scutiformis (Teller), M. ochotica densistriata (Teller), M. ochotica ochotica (Keyserling) and M. sabaikalica (Kiparisova) with M. mabara (Kobayashi and Ichikawa) (Tables 2, 3).
|