CHAPTER 8
Geographic Variation of Cryptopecten vesiculosus
|
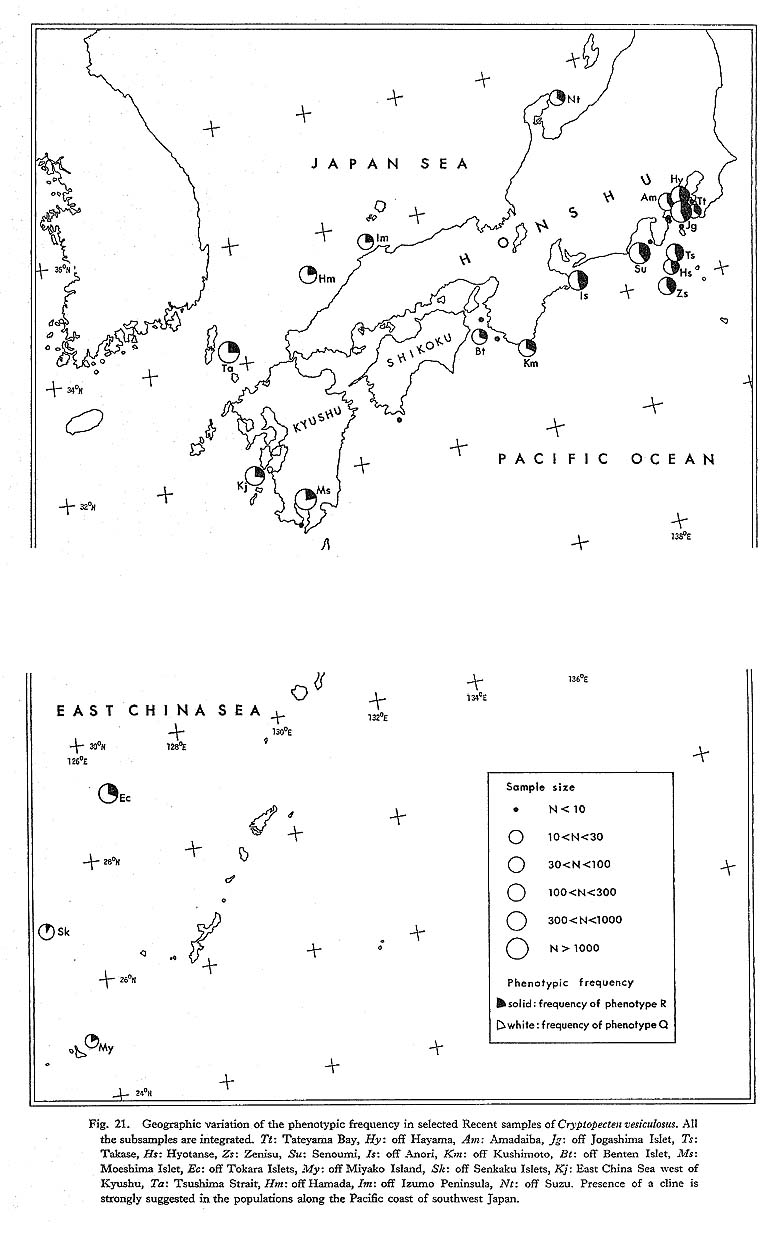
Geographic variation is, so to speak, spatial integration of intrapopulational variation, and evolutionary change within a lineage is chronological integration of geographic variation. Quantitative examination of geographic variation seems to be indispensable for the study of phyletic evolution as well as speciation. Gould and Johnson (1972) emphasized the importance of the study of geographic variation and the potential role of multivariate analyses which can treat the variation of many characters (morphological, physiological, biochemical, etc.) simultaneously. In the present article, however, I hesitate to apply this method to the morphometric data of Cryptopecten, chiefly because causal evaluation of various unit characters is insufficient. Some characters may represent genetic difference between populations, while other characters may be strongly influenced by environmental factors. Various kinds of heterogeneity among samples also precludes this approach. Consequently, I intend to clarify the morphological differences among population samples on the basis of each univariate or bivariate character and then to approach synthetically the general aspect of geographic variation. Ideally, geographic variation should be determined from many large population samples covering the whole distribution of the species in question. Among the three living species of Cryptopecten, reliable conclusions may be obtained only for C. vesiculosus, whose distribution is virtually restricted to the seas around the central and southwestern part of the Japanese Islands. Some qualitative observations were made also on the geographic variations of C. bullatus and C. nux, which distribute more extensively than C. vesiculosus, and the results will be shown in the section of systematic description. In spite of recent progress in the chronology of Neogene and Quaternary marine deposits in Japan, the geographic variation of C. vesiculosus in the geological ages is still difficult to clarify. More than 15 large Recent samples of this pectinid, including more than 100 subsamples, are available for the study of geographic variation, though the material is still not necessarily ideal. The localities of large samples concentrate on the Pacific coast of central Honshu (Kanto and Tokai Regions) and the western coast of Kyushu, and are rather sparsely situated in the Japan Sea and the seas surrounding the Ryukyu Islands. The well-preserved large Holocene fossil sample Ms (only about 2,720 years B. P. according to Matsushima's 14C dating) from south Kyushu may also be useful for the study of geographic variation. 1. Shell Size and Growth Rate [Table 13]
Generally speaking, body size is regarded as an easily measurable character that varies geographically. In the present case, however, there are some problems, because the age (and size) composition of the samples may be quite variable. The height of the maximum individual in each sample ( On the basis of analysis of several large Recent samples of C. vesicitlosus, it can be said that the geographic variation of size, represented by the difference of the values of 2. Shape of Shell (Bivariate morphometric characters) [see Tables 4-10]
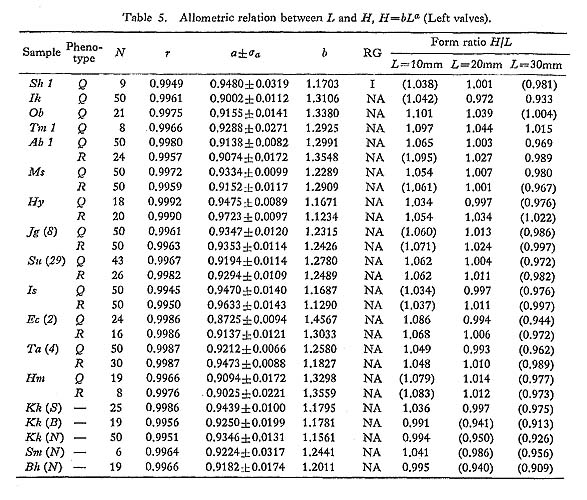
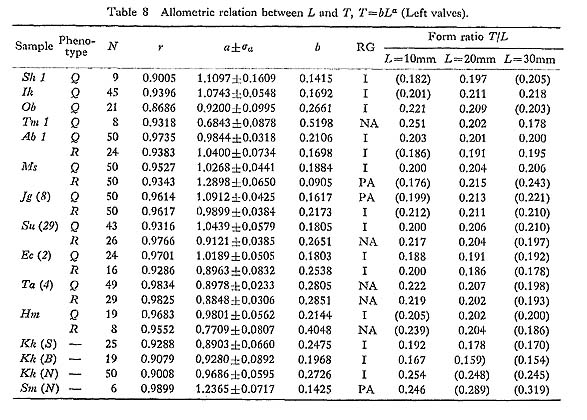
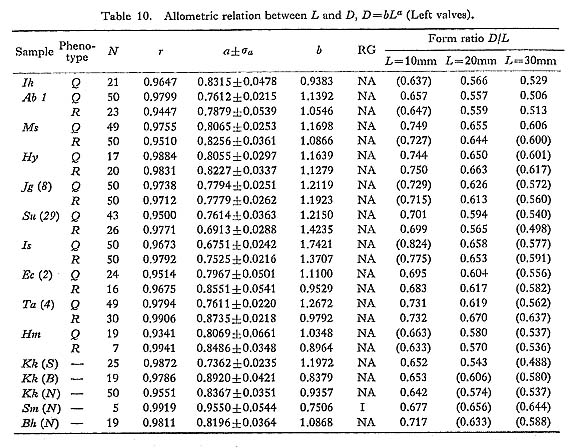
The results of various bivariate analyses on seven Recent samples and one Holocene fossil sample seem to offer data for the consideration of geographic variation. The allo metric relations between L and H, L and T, and L and D as well as standardized form ratio H/L, T/L and D/L in these samples were compared. Although these bivariate characters sometimes differ significantly between samples, no conspicuous tendency of geographic variation is detected here. 3. Number of Radial Ribs [see Tables 11 and 14]
As mentioned before, the frequency distribution of the number of radial ribs on the disk (X) is unimodal and essentially normal in every sample, the mode lying at 14 or 15 in all Recent samples. However, the mean
4. Coloration of ShellThe geographic variation of color pattern and ground color is intuitively not much different among Recent samples. Every large sample contains yellow-orange monotonic individuals and shows distinct color polymorphism. The phenotypic frequency is, however, commonly less than 5 percent and too unreliable to be applied as an indicator of geographic variation. 5. Surface Sculpture (Phenotypic frequency) [see Tables 12 and 15]
The two discrete phenotypes, Phenotypes Q and R, are clearly separable in every Recent sample, and no individual is intermediate. The relative frequency of Phenotype R as well as a range of 95 percent confidence (obtained by small sample method) was calculated in each sample on the basis of all the constituent individuals. In the previous study (Hayami, 1973), I concluded by means of chi-square tests that no significant difference of phenotypic frequency exists between any pair of Recent samples, and that the living populations of C. vesiculosus may be considerably panmictic. The samples treated at that time, however, are from only a limited sea area, the Pacific coast of central Honshu. Also I was uneasy about the variation of the Holocene fossil sample Ms, whose phenotypic frequency is unexpectedly low for its very young geological age. As a result of the present study on samples from much wider sea areas, it was found that phenotypic frequency is often significantly different between samples of distant areas, particularly between samples from the Pacific coast of central Honshu and samples from the East China Sea-Japan Sea side of the Japanese Islands, as documented by a newly prepared chi-square matrix (Table 15). All the large samples from the Pacific coast of central Honshu (e. g., Hy, Jg, Am, Zs, Ts, Su and Is) show nearly the same phenotypic frequency within a range from 0.39 to 0.46, whereas in the samples from the East China Sea and Japan Sea (e. g., Sk, My, EC, Kj, Ta, Hm and Im) the frequency is much lower, ranging from 0.09 to 0.30. Therefore, the Holocene fossil sample Ms from south Kyushu is not in the least unusual with respect to this character. The samples Km and Bt off Kii Peninsula appear to be intermediate between the two clusters as is their geographic location (Fig. 21).
The geographic variation of this character also indicates the presence of an evident cline along the Pacific coast of the Japanese Islands. The available data for the East China Sea and Japan Sea do not show any conspicuous geographic variation. However, the northernmost sample Nt off Noto Peninsula, though of relatively small size, shows considerably high phenotypic frequency, and therefore it is probable that somewhat similar clinal change also exists along the Japan Sea side. It is remarkable that the conclusion on the geographic variation of this character roughly coincides with that on the average number of radial ribs. 6. Causal Evaluation of Geographic VariationThe genetic background for the variation of these characters is still unknown. As it dwells only in environments under the influence of oceanic water, Mendelian experiments on this pectinid are probably more difficult than on molluscs in coastal waters. Circumstantial evidence about the intrapopulational and geographic variations, however, convinces me that some of these unit characters are strongly controlled by genetic factors. Among others, the number of radial ribs on the disk and the relative frequency of phenotype R, both of which show obviously clinal changes at least along the Pacific side of the southwestern part of the Japanese Islands, are most striking. The number of ribs is probably polygenic in view of its normal-type frequency distribution and uniformity of variability in each sample. The dimorphism is, on the other hand, probably determined by a single (or a few) genetic factor such as an allele or chromosomal aberration, as I presumed somewhat boldly before (Hayami, 1973). As discussed by Mayr (1963, p. 361), dines are generally the products of two conflicting forces: one is the differential selection pressure in accordance with local environments (or environmental gradient), and the other is gene flow among local populations. The former acts to increase the difference of gene frequency among local populations, while the latter contributes to homogenizing the gene frequency throughout the distribution of the species in question. Because the present pectinid is an open sea dweller, a considerable amount of gene flow can be expected during planktic larval stages. Water temperature evidently decreases from southwest to northeast on both sides of the Japanese Islands and may be one of the significant factors causing the environmental gradient. At least on the Pacific side, the clinal change of phenotypic frequency as well as average number of radial ribs seems to correlate well with the gradient of water temperature. It is, however, difficult to say what environmental factors are actually responsible for the differential selection pressure, because, as pointed out by several scientists, significant correlation does not necessarily mean direct causal relation. |
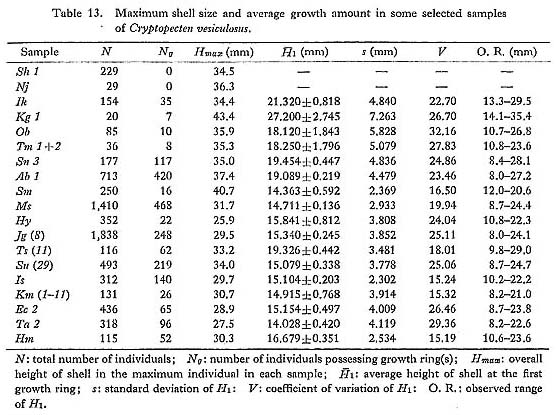
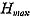 ) may be informative, but it is certainly influenced by sample size. In this respect the average size (height) at the first stepwise growth ring (
) may be informative, but it is certainly influenced by sample size. In this respect the average size (height) at the first stepwise growth ring (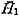 ) may be more meaningful, although the intrapopulational variation of this character is generally wide.
) may be more meaningful, although the intrapopulational variation of this character is generally wide.




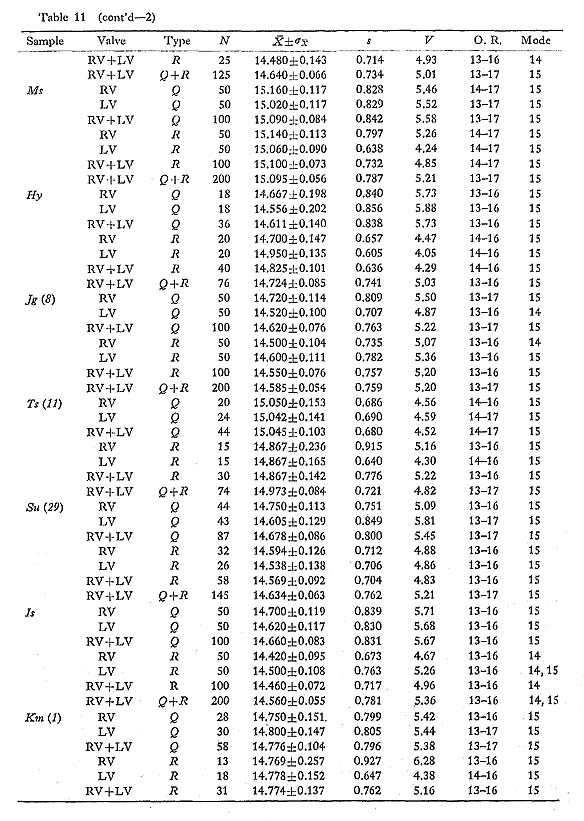

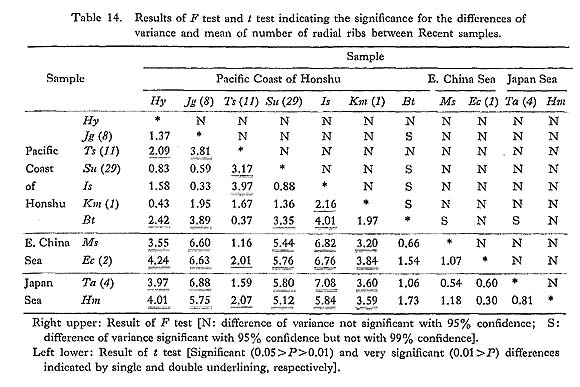
 is often significantly different between samples of different sea areas. It is as small as 14.5-14.8 in the samples from the Pacific coast of central Honshu, while it is commonly larger than 15.0 in the samples from the East China Sea and Japan Sea (Fig. 20). The Holocene fossil sample Ms from south Kyushu seems to belong to the latter cluster in this character. The sample Ts (11) and some other small samples from three sea banks off the Izu-shichitô Islands, and the sample Bt from Kii Strait between Kii Peninsula and Shikoku are actually intermediate between the two clusters. The results of F test and t test are collectively shown in Table 14. These data seem to indicate that the geographic variation of this character constitutes a gentle dine along the Pacific coast of the Japanese Islands; the average number of radial ribs becomes more numerous towards the warm southwest seas. This clinical change, however, is not apparent along the coasts of the East China Sea and the Japan Sea.
is often significantly different between samples of different sea areas. It is as small as 14.5-14.8 in the samples from the Pacific coast of central Honshu, while it is commonly larger than 15.0 in the samples from the East China Sea and Japan Sea (Fig. 20). The Holocene fossil sample Ms from south Kyushu seems to belong to the latter cluster in this character. The sample Ts (11) and some other small samples from three sea banks off the Izu-shichitô Islands, and the sample Bt from Kii Strait between Kii Peninsula and Shikoku are actually intermediate between the two clusters. The results of F test and t test are collectively shown in Table 14. These data seem to indicate that the geographic variation of this character constitutes a gentle dine along the Pacific coast of the Japanese Islands; the average number of radial ribs becomes more numerous towards the warm southwest seas. This clinical change, however, is not apparent along the coasts of the East China Sea and the Japan Sea.