





Boniniella ikenoi (Makino) Hayata
シダ植物`ャセンシダ科
東京都小笠原諸島母島石門山
服部広太郎
1905年8月9日採集
理学部附属植物園
私は、研究対象としては学生時代からホウビシダ類というチャセンシダ科の一群を研究してきた。結果的に卒業研究の時から、もう14年間もホウビシダ類の研究をやってきたことになる。一方、研究手法は様々なものを用い、形態や生態はもとより、生殖様式や染色体などの細胞学的形質、化学成分、果てはDNAまで様々な角度からホウビシダを調べてきた。同時に研究対象地域も広げて、中国や、東南アジア、インドシナ、中南米など世界中のいろいろな場所に行って、地球レベルでホウビシダ類の研究をしてきた。今回は、その成果のあらましをご紹介したいと思う。
本稿では、まずどのような種がホウビシダ類にあるか、という話をする。広義のホウビシダ(Aspleni umunilaterale)は用不用説で有名なラマルクによって記載された種である。そんなに昔から知られているならホウビシダ類の種の認識ぐらいは、私が研究を始める前に大方わかっていたんだろうと思われるかも知れないが、そうではない。種の認識の仕方を論理的なものに改良することによってホウビシダ類の種数はすでに3倍近くになった。私の研究が進めば、ホウビシダ類の種数はまだまだ増えることだろう。
次に、それらの種の間の系統関係がどのようになっているか、という話をする。私は、形態や化学成分に始まり、最近ではDNAとこれまた様々な形質情報を用いて系統解析をしてきた。種が認識できたなら、それらの間の系統解析をすることが必須である。というのは、系統樹を得ることは、それぞれの種の進化の歴史を理解することにつながるからである。
さらに系統樹が得られれば、それをもとにしてホウビシダのもつ興味深い形質の進化を科学的・論理的に議論することもできる。そこで、系統解析とともに形質進化の話も一緒に紹介したいと思う。
ホウビシダ類は世界中の熱帯に広く分布している。その中でもホウビシダ(広義)という一つの種は、私が研究を始めた時点では、アフリカから、アジア、ハワイまで旧世界の熱帯中に広く分布しているということになっていた。ところが、ホウビシダは日本にも分布しているが、日本のものだけを見ても形態的にも細胞学的にも変異が大きいことが知られていた。この細胞学的に変異が大きいというのは、ホウビシダでは通常の有性生殖をするもののほかに無配生殖をするものがあることが報告されていたのである。無配生殖とは減数することなく複相の胞子が形成され、これから生じた複相の前葉体の一部から接合なしに次世代の胞子体ができるという生活環をもつようになったもののことをいう。減数と接合をともに失った(要するに性を失った)ことで、無配生殖をするものでは子供はすべて親と遺伝的に同一なクローンとなる。このような無配生殖はシダ植物では、それほど稀な現象ではなく、日本産シダ植物の17パーセントが無配生殖種である。
「日本のホウビシダには、無配生殖をするものと有性生殖をするものがあるから、これら2つの生殖型が地理的、形態学的にどのようになっているかを調べてみなさい」というのが岩槻邦男教授から与えられた私の卒業研究のテーマだった。シダ植物では胞子嚢あたりの胞子数を数えることで、さく葉標本でも簡単にその生殖様式を推定することができる。有性生殖をするものでは、1つの胞子嚢に64個の胞子が入っているのに対して、無配生殖をするものでは半数の32個入っているからである。ところで、テーマをくださった岩槻先生の予想では形態的には2つの生殖型の間で差がなく、地理的にもいろいろな場所で無配生殖が見られ、ホウビシダという一つの種の中で無配生殖が何回も進化していることを実証できるのではないかということであった。
ところが、実際に調べてみるとこの予想は外れて、2つの生殖型は地理的に分かれて分布していた。すなわち、関東から九州中部にかけては無配生殖型だけが、九州南部から沖縄にかけては有性生殖型だけが、そして、沖縄以南ではまた無配生殖型が分布している(挿図1)ことがわかった。さらに、この日本本土に分布する無配生殖型、有性生殖型および、沖縄以南に分布する無配生殖型の間には形態的にも差があることがわかった。
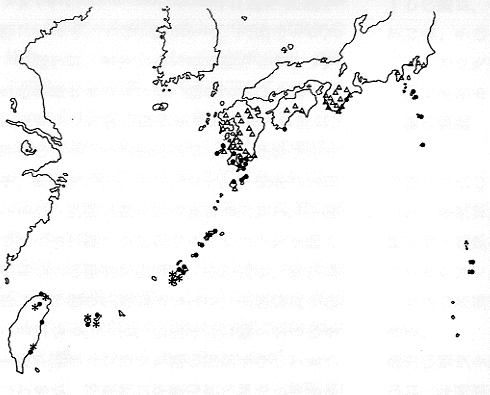 | 14-1 日本産ホウビシダ3型の地理的分布。△:日本本土に分布する無配生殖型、●:有性生殖型、*:沖縄以南に分布する無配生殖型。 |
日本本土に分布する無配生殖型は羽片が平行四辺形で胞子嚢群が羽片の縁から離れてつくのに対して、有性生殖型では羽片は鎌形で、胞子嚢群は縁によって付く。沖縄以南に分布する無配生殖型では羽片の先が丸く、根茎も前二者と比べて短いという違いもあることがわかった。さらに、例外的に日本本土に分布する無配生殖型と有性生殖型が同所的に分布する紀伊半島の尾鷲、および、有性生殖型と沖縄以南に分布する無配生殖型が共存する石垣島のバンナ岳で野外調査をした。その結果これら3型がはっきり区別できるものであることが確信できた。というわけで、この2つの無配生殖種は新種として記載した。
この研究を通して私は自分のように野生植物に対する素養がなくても、これまでの分類学者がもっていなかった観点をしっかりもって比較研究すれば、新しい種を認識することもできることを知った。また、同所的に複数の種が分布しているようなフィールドで、生育環境まで含めて比較観察することがホウビシダ類のように形態が単純な植物の種を認識する上で役に立つことも知った。さらに、ホウビシダは一つの種が旧世界に広く分布していることになっていたわけであるが、日本のものだけでも3種が存在したことがわかったわけである。世界のその他の地域のものも、日本産のものと同等の精度で調べれば、たくさんの種があるであろうことは容易に想像がつく。
というわけで、以後、世界の熱帯地域の中でもホウビシダ類の多様性が特に高い中国西南部、東南アジア、中南米などに自ら赴いて、(1)同所的に分布していて、なおかつ区別できるものは別種である。(2)種が違えば生育環境も異なることが多い。以上の2点を念頭において新しいホウビシダ類の種の認識に努めてきた。これから、中南米と中国での調査を例に上げて、私の種の認識の実際を紹介してみたいと思う。
(2)中南米産ホウビシダ類の分類
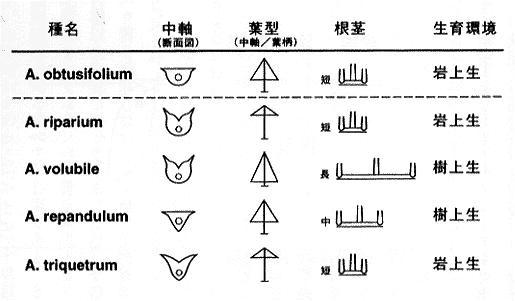
アメリカのミズーリ植物園に1年半留学したときに、新世界のホウビシダ類を研究した。私が研究をする以前には新世界のホウビシダ類のモノグラフはなかったが、中南米各国の植物誌を見ると、アメリカのシダ学者は5種ないし6種を認識していた。研究者間で意見の不一致があったのは、A. obtusifolium とA. repandulumが区別できるものか、それとも変異が連続した一つの種かということであった。私の研究の結論から先にいうと、この2種は別種で、さらに A. repandulum(広義)は四種に分けられるというものである。その結論に至った過程を次に説明する。
パナマのフォルツナというところにはA. obtusifoliumとA. repandulum(広義)が同所的に存在していた。A. obtusifoliumは渓流の常に水しぶきがかかるようなほとんど半水生といってよい環境に生えている。葉はワカメやコンブのように透き通っている。日本のヤクシマホウビシダとちょうど同じ生育環境(渓流沿いの常に水しぶきのかかる岩上)で、葉が透き通っているのもヤクシマホウビシダと同じである。一方、A. repandulumの生育環境はやはり湿った渓流沿いの岩の上であるが、A. obtusifolium よりも水際から離れて水しぶきはかからないような環境に生育している。葉も透き通ってはいない。
挿図2は葉の断面を示したものである。A. repandulum(広義)は普通のシダ同様、細胞層は7〜8層からなり、細胞間隙も良く発達している。それに対して、A. obtusifoliumの葉は細胞層が4層と薄く、細胞間隙もほとんど発達していない。これが、葉が透き通って見える理由である。 A. obtusifolium(広義)とA. repandulum(広義)では葉に違いがあるだけでなく、たとえば胞子の表面模様にも違いがあることがわかった。他のホウビシダ類も含めてチャセンシダ科のシダは、一般にひだが良く発達した胞子をもっている。A. obtusifoliumと、広義の A. repandulumはともに、ひだのない特殊な胞子をもっているが、さらにこの両者では違いがある。広義のA. repandulumでは、長い刺が多数あるような胞子であるのに対して、A. obtusifoliumでは短いこぶ状の突起がある胞子をもっている。このように、A. obtusifoliumとA. repandulum(広義)の間にはいろいろ違いがある(挿図3)。
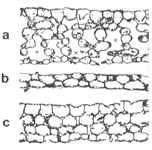 |  |
| 14-2 中南米産 A. obtusifolium 群の分類。 | 14-3 ホウビシダ類の葉の断面。a)ホウビシダ類一般、b)ヤクシマホウビシダ、c)A.obtusifolium |
一方、A. repandulum(広義)はいくつかの種に分けられるということを先に述べたが、パナマのフォルツナには、岩の上に生えるもの(狭義のA. repandulumと区別して、A. ripariumと呼んだ)の他に、細い木に根茎を蔓状に巻き付けて生えるものもあった。これは後にA. volubileと名付けた。A. ripariumとA. volubileは、根茎の長さの他に葉型も少しずつ異なり、現地で一緒に生えているところを観察すれば容易に別種と認識できるものだと思うが、当時誰もこの違いを認識していなかった。以上、フォルツナにあったA. obtusifolium、A. riparium、A. volubileの3種は、同所的に存在するものの、生育環境が少しずつ異なり、また形態学的にも不連続性が観察された。
さて話は続いて、今度はエクアドルに行ってみると、先ほどあった、細い木にぐるぐる巻き付くA. volubileに加え、太い木の幹に短く根茎をはわせて生えるものがでてきた。この両者には葉の形態にも差が見られた。A. volubileの葉は上から見ると、中軸の部分に縦に翼が発達して溝がある。そして、葉の裏側を見ると、中軸は丸いかたちをしている。それに対して、太い木の幹に付くものでは中軸の表側は翼が水平に発達するため平らである。中軸の裏側は断面で見ると三角形になっている。このように、一度わかれば違いが次々見つかって、木の上に生える種も2つあることがわかった。
以上のような観察を繰り返すことで中南米のA. obtusifoliumとA. repandulum(広義) は中軸の構造、葉型、胞子の表面模様、根茎の長さおよび生育環境で区別できる五つの種類に整理することができた(挿図3)。ホウビシダ類のように形態学的な変化がそれほど大きくならないような群でどのような種が存在するかを知るには、このように同所的に分布するものの詳しい比較観察をするのが近道である。
(3)中国産ホウビシダ類の多様性
同様の研究は、たとえば中国のシーサンパンナでも行った。その結果、ここでもホウビシダ類の種数は3種から7種に増えた。さて今度はこの研究を例にあげて、種を認識する研究はただそれだけにとどまらず、その生物群のもつ形質の多様性の発見にも貢献することをお話ししたい。
1つめの例は、私たちがシーサンパンナで発見したミドリラハオシダモドキという変種である。これは一見すると東南アジアにあるミドリラハオシダとよく似ているが、注意深く観察すると胞子嚢群がソーセージ状で、そもそもチャセンシダ科に共通の胞子嚢群のかたちと違っている。ホウビシダ類も含めチャセンシダ科のシダは700種もあるが、ほとんど例外なく、この挿図4aのように1枚の包膜で覆われた胞子嚢群をもっている。それに対して、ミドリラハオシダモドキでは挿図4bのように2枚の包膜で挟まれたような包膜をもっていた。胞子嚢群は種子植物では花に相当するもので、この包膜の構造の違いは昔なら新しい科をたてただろうというぐらいの大きな違いである。もちろん、私は包膜の形の違いだけで新しい科を造ったりはしない。しかし、科の定義に使えるほど、長い進化の歴史においても安定している形質が変化しているものを見出したことは興味深い。
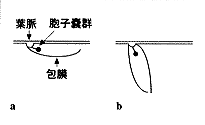 | 14-4 ミドリラハオシダモドキの包膜 |
次にもう一つ、中国での種を認識しようとする作業の過程で、日本にあるホウビシダ類の起源に深く関わったと思われるものを見出した例を紹介する。最初に紹介した生殖様式に着目することで私が新たに認識した沖縄以南に分布する無配生殖種は現在ではタイワンホウビシダと呼んでいるが、この種は後に詳細な細胞分類学的研究を行っても、沖縄から台湾にかけての地域には3倍体無配生殖型しか見出せなかった。ところが、中国のシーサンパンナでは、3倍体無配生殖型に加え、少なくとも外部形態的には、これとほとんど変わらない2倍体有性生殖型、さらに4倍体有性生殖型のタイワンホウビシダも見出された。無配生殖型がどのようなメカニズムで生じるかは、まだよくわかっていないが、これらが有性生殖型から生じたことは、ほぼ間違いないであろう。したがって、日本産の無配生殖種のもとになったものを中国で見つけたといってもいい過ぎではないと思う。このように、外国で種を認識するための研究は日本に分布する種の起源に関わった原始的な植物種を探すことにもつながるのである。
(1)解剖学的解析
ホウビシダ類はチャセンシダ科にあって特異な背腹性のある横に長くはう根茎をもっている。背腹性があるというのは、根茎の上面から葉が出て、下面から根が出ることをいう。他のチャセンシダ科の種は、ほとんど例外なく葉がらせん状につく短い根茎をもっている。早田文蔵博士は根茎の柔組織をピンセットと針で取り除き、維管束だけを残すことによって、その構造を直接観察する方法を開発した。そしてこの方法を用いて数多くのシダ植物の根茎中の維管束構造を観察した。ホウビシダでは、上下に背腹2本の太い維管束が走り(挿図5)、これらから1本ずつ合計2本の葉跡(葉にはいる維管束)が出る。そして葉跡の反対側で上下の維管束が連結されることで葉隙を形成する構造が見られることを早田博士は明らかにした。その後、岩槻博士と加藤雅啓博士(1975)は早田の方法を用いて当時認識されていたホウビシダ類のすべての種について維管束構造を調べ、ホウビシダの近縁種群はすべてホウビシダと同様の維管束構造をもつことを明らかにした。
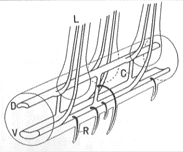 | 14-5 ホウビシダの根茎中の維管束構造。D)背側の維管束、V)腹側の維管束、L)葉跡、C)連結脈、R)根跡。 |
さて、ホウビシダ類とよく似た背腹性のある匍匐根茎をもつチャセンシダ科の種(As-plenium obtusifolium群)が中南米にもあることは昔から知られていた。しかしこれらが 旧世界のホウビシダ類と直接系統的なつながりがあるものかどうかは不明であった。そこで私は、自分で認識できたA. obtusifolium群すべての種について、やはり早田の方法により根茎の解剖を行った。その結果、A. obtusifolium群もアジアのホウビシダ類と全く同一の維管束構造をもつことがわかった。このことは、新旧世界を問わず背腹性の匍匐根茎をもつチャセンシダ科の群は、系統的にも近いことを強く示唆している。この結果は、後のrbcLの塩基配列比較に基づく分子系統学的解析によっても支持された。
(2)植物化学的解析
私は大学院の修士の頃は、もっぱら天然物有機化学をやっていた。具体的にいうとホウビシダ類から化学成分を単離して、質量スペクトルや核磁気共鳴スペクトルによってその分子構造を解析・決定するというものである。そしてホウビシダ類がとても変った酸性のアミノ酸を複数もっていることを明らかにした。ホウビシダ類は、グルタミン酸よりも炭素鎖の2つ長くなった2-アミノピメリン酸およびその4-位が水酸基によって置換されたもの、さらに不飽和化されて二重結合が入ったものなどを植物体内に大量に蓄積していることがわかった(挿図6)。以後、これらのアミノ酸をそれぞれ、APA、OH-APA、Δ-APAと呼ぶことにする。このような、タンパク質を構成しないアミノ酸は生物界から七百種以上も見つかっている。ただしその立体配置についてはタンパク質を構成するアミノ酸同様ほとんどすべてがL型である。ところが、ホウビシダのΔ-APAとAPAは自然界からは 非常に珍しいD型の立体配置をもつアミノ酸であることもわかった。このように特異な物質の分布は系統を反映することが期待できるので日本産ホウビシダ類におけるこれらのアミノ酸の分布を調べてみた。その結果、APAとOH-APAはすべての種がもっていたが、Δ-APAについてこれをもつものともたないものがあることがわかった。さらに形態と Δ-APA の分布パターンを比較してみると、Δ-APAをもつ種は根茎が長くはい、根は細く、さらに根は根茎の下面だけから出るのに対して、Δ-APAをもたないものは、根茎が短くはい、根は太く、さらに根は根茎の上面からも出ることがわかった。外群として用いたウスバクジャクは、Δ-APAをもたず、根茎の形態も後者の方と一致した。
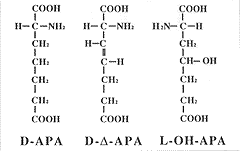 | 14-6 ホウビシダ類のもつ特異なアミノ酸。APA、2-アミノピメリン酸、Δ-APA、3、4-デヒドロ-2-アミノピメリン酸、OH-APA、4-ヒドロキシ-2-アミノピメリン酸。 |
以上の結果を分岐分類学的に総合すると、Δ-APAをもつ3種が系統的に近縁であることになる。この研究を行った当時、私はそう結論づけた。とはいえ、本当のことをいうと、これらの根茎に関わるいくつかの形質が互いに独立である保証はない。また、これらの形質が適応的に何回も平行進化したことも十分ありそうである。実際、後で紹介するrbcLの塩基配列比較による分子系統学的解析の結果では、Δ-APAをもつ3種が系統的に近縁であることは支持されなかった。
(3)細胞学的解析
染色体数については、チャセンシダ科の200以上の種について報告がある。そしてホウビシダ類以外はほとんど例外なくx=36の基本数に基づく染色体数である。したがって、染色体の基本数が36であることは、チャセンシダ科を特徴づけるよい形質の一つとなっている。ところがホウビシダ類では1954年のマントン(I. Manton)とスレッジ(W. A. Sledge)のx=40の報告に始まり、x=36'、38'、40'と様々な基本数に基づくと考えられる染色体数が報告されていた。そこで、三井邦男博士、岩槻博士と私は日本産のホウビシダ類を中心に減数分裂と体細胞分裂の両方を観察することで正確な染色体数の決定を行った。また、この両方を観察することは胞子嚢あたりの胞子数から間接的に推定していた無配生殖の確認にもつながる。なぜなら、有性生殖を行うものでは、減数分裂時の染色体数(n)が体細胞分裂時のもの(2n)の半分になるのに対して、無配生殖を行うものではこれらが同数になるはずだからである。
結果を表1に示した。これを見ると、ホウビシダ類の染色体の基本数は、今までの報告とは異なりx=39であることがわかる。ウスイロホウビシダだけがx=38であった。これらの染色体の基本数は他のチャセンシダ科のものとは異なっており、背腹性の匍匐根茎とあわせてホウビシダ類を特徴づけるよい形質である。また、胞子数の計数から無配生殖と推定していたものは、確かにそうであることも確認できた。
 | 表1 ホウビシダ類の染色体数。n:滅数分裂時の染色体数、2n:体細胞分裂時の染色体数、——:未観察。 |
(4)分子系統学的解析
近年DNAの塩基配列レベルの情報が分子生物学的技術の進歩により、野生植物からも容易に得られるようになった。特定の遺伝子の試験管内高速コピー機であるPCR法と自動DNA塩基配列決定装置(DNAシークエンサー)によるダイレクトシークエンス法を組み合わせれば、2.5日で1つの野生植物の1200塩基対程度の配列を決定できる。私たちは、rbcL遺伝子の塩基配列を直接決定・比較することによってホウビシダ類の系統解析を試みた。rbcL遺伝子はリブロースビスリン酸カルボキシラーゼという光合成の際に炭酸固定を司るとても重要な酵素タンパク質の大サブユニットをコードしている。これはアミノ酸レベルでは非常に保存的で、陸上植物の中では系統的に離れたシダ植物と種子植物のものを比べても20パーセント程度しか異ならない。そこでこれまで、科以上の高次分類群間の系統解析に用いられることが多かったようである。しかし、塩基レベルでは変化の早い部位もたくさんあるので種間の系統解析にも十分有効である。
挿図7は、rbcLの塩基配列の比較に基づいて最大節約法によって推定された日本、中国および中南米産ホウビシダ類の分子系統樹である。枝の上の数字は1万回のブートストラップを行った際の各枝の再現性をパーセント表示したものである。もちろん再現性が高いほど信頼性も高い。これを見ていただければわかるように、解像度、信頼性ともに高い系統樹が得られている。挿図7によると、アジア産の種は単系統群であるが、中南米産の種には大きく2つの群(A. obtusifolium群とA. laetum群)が存在し、これらは単系統群ではないかも知れない(ブートストラップ確率は60パーセントと低い)ことがわかる。
ホウビシダ類がどのようなものかを知るためには、ホウビシダ類だけを調べているだけでは不足である。ホウビシダ類がチャセンシダ科に属することは、ほとんど疑いがないので、ホウビシダ類に加えてチャセンシダ科の様々な種もいれてrbcLの比較解析をした結果が挿図8である。分子系統学の利点の一つは非常に近縁なものから結構遠縁のものまで一緒に混ぜて系統解析できるところにある。ホウビシダ類は従来、葉型の似たヌリトラノオなどから比較的最近、特殊化して生じた群と考えられてきた。ところが、挿図8を見ると、ホウビシダ類はチャセンシダ科の多様化の一番初期に他の種群から分化した起源の古い単系統群であることがわかる。別のいい方をすると、チャセンシダ科の他のどれとも近縁でないということになる。ホウビシダ類は、たとえばホウビシダ属として別属で取り扱ってもよいことも、この分子系統学的解析からわかった。
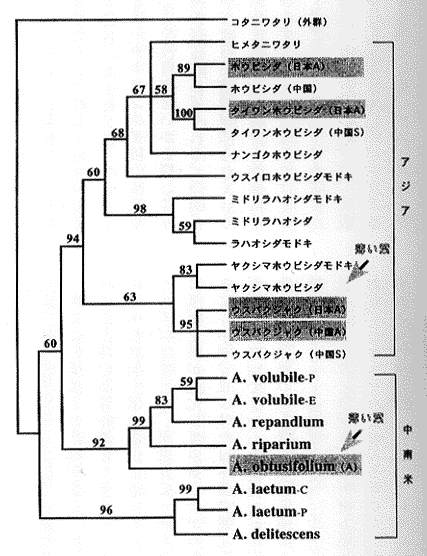 | 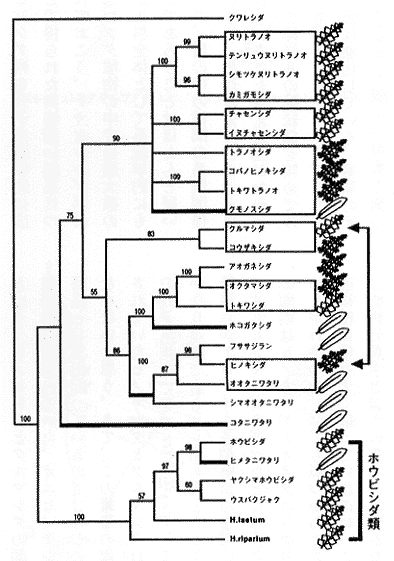 |
| 14-7 ホウビシダ類のrbcL遺伝子の塩基配列に基づく分子系統樹。網部分は無配生殖をする種。 | 14-8 チャセンシダ科の分子系統樹。右端に葉形の模式図を示す。単葉が進化した枝は太線で示す。 |
ここでチャセンシダ科を分子系統学的に広く解析することで得られた興味深い結果についても少し触れておこう。チャセンシダ科は700種以上を含むシダ植物の中では最大級の科である。昔から科全体として系統学的にもまとまっているであろうことはほとんど疑いがなかったものの、科の中の種間の系統関係はさっぱりわからず、それゆえにどのように属をもうけるかということさえ定説がなかった群である。さて挿図8の系統樹を眺めてみると、たとえば単葉は、チャセンシダ科の中で少なくとも4回は進化している。単葉のオオタニワタリ(挿図9a)と細かい複葉で葉の先に無性芽をつけるヒノキシダ(挿図9b)は今回の分子系統学的研究によって非常に近縁であることが示されたが、これらは外見上は何も似ていない。このように葉形はチャセンシダ科の系統をほとんど反映していないのである。 それでは、なにかチャセンシダ科の系統を反映しているような性質がないかと探してみると、自然雑種の存在があげられる。挿図8では、そのあいだで自然雑種が知られている種群の種名を四角で囲った。ふつう雑種は系統の遠近に関わらず、たまたま近くにはえると偶然生じるものと考えられている。しかし、少なくともチャセンシダの中では、比較的近縁な種間でしかできていないようである。これに反する例として、オニヒノキシダという、ヒノキシダとクルマシダの間の推定雑種が報告されていた。オニヒノキシダは園芸品として出回っているのでご存じの方も少なくないと思う。さて、この雑種の両親種とされていたヒノキシダとクルマシダは挿図8の分子系統樹で見ると(矢印で示してある)、かなり遠縁であることがわかる。ところが、筆者らがアロザイム多型解析法を用いて、このオニヒノキシダを調べたところ、この雑種の両親はヒノキシダとオオタニワタリであることがわかった。似ても似つかぬこの2つの近縁種は雑種まで造っていたのに、これまでの植物分類学者はこの事実に気付かなかったというわけである。このことは、分子系統学的手法の威力の大きさも同時に示している。
やはり中国で見つけた特殊な胞子嚢群(包膜が2枚)をもつミドリラハオシダモドキは胞子嚢群以外の形態が良く似たミドリラハオシダと近縁であることがわかった。このことは逆に、特殊な胞子嚢群のかたちが、ミドリラハオシダモドキでは短期間に進化したことを示している。七百種もあるチャセンシダ科の中で安定している(それゆえ変わりにくい)包膜の構造がなぜミドリラハオシダモドキで変化し得たかという発生学的メカニズムの解明は今後の興味深い研究課題である。
同様の例は、ヒメタニワタリでも見られる。ヒメタニワタリは、小笠原の母島、沖縄の北大東島、および中国の海南島の3ヶ所だけに隔離分布する種で、挿図10bのようにスペード型の特殊な形の葉をもっている。少なくとも葉型だけを見るとホウビシダ(挿図10a)とは似ても似つかない。しかし、ホウビシダ類同様、背腹性のある長くはう根茎をもつ。かつて、早田博士は、これらの形質に着目してヒメタニワタリ属(Boniniella)をたてたりもした。加藤博士ら(1990)は、このヒメタニワタリの根茎中の維管束構造および体細胞染色体の数を調べた。その結果、維管束構造はホウビシダ類と同一であり、染色体数もn=78で、基本数はやはりホウビシダ類と同じく39であることがわかった。これらの結果はヒメタニワタリがホウビシダ類の一員であることを強く示唆している。今回得られた分子系統樹(挿図7、8)によってもヒメタニワタリがホウビシダ類に類縁のあることは支持された。さらに驚いたことには、ヒメタニワタリはホウビシダ類の多様化の歴史の中で初期に他の種から分化したのではなく、比較的最近にナンゴクホウビシダやホウビシダ(狭義)の群から進化したこともわかった。このことは葉型が短時間で大きく変化することを示している。
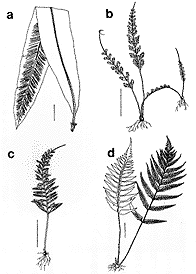 | 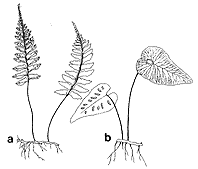 | |
| 14-9 a)オオタニワタリ、b)ヒノキシダ、c)オニヒノキシダ、d)クルマシダ | 14-10 a)ホウビシダ、b)ヒメタニワタリ |
先に、アジア産のヤクシマホウビシダと中南米産のA. obtusifoliumが半水生の特殊な生育環境と細胞間隙の消失した特殊な薄い葉をもつことを紹介した。このように特定の生物が特異な生育環境と特殊化した形態形質を両方もっていると、それだけでこの両者の間に因果関係をつけて、薄い葉は半水生の生育環境に適応して進化したといった具合に考えてしまいがちである。しかし、これは論理的には正しくない。たとえば、ヤクシマホウビシダとA. obtusifoliumがごく近縁だとすると、これら2つの性質を共通の祖先で1回獲得すればよいことになる。過去に1回だけ起こったことに必然性(起こった理由)が存在するとは限らないので、これでは薄い葉の進化的意義を考えても科学的には無意味である。ところが、挿図7の系統樹を見てみると、ヤクシマホウビシダとA. obtusifoliumはホウビシダ類の中では比較的遠縁である。しかもそれぞれ、ウスバクジャクやA. ripariumといった通常の陸上生の生育環境と細胞間隙も発達した種と近縁である。ホウビシダ類の中では半水生の生育環境と特殊な薄い葉はそれぞれ2回独立に進化したことになる。したがって、半水生の生育環境に適応して細胞間隙のない薄い葉が進化したと議論することが可能になるのである。このように系統関係を解明することで、ホウビシダ類のもつ興味深い形質の進化を科学的に議論することができたのである。
もう一つこれからやろうと思っているのは、ホウビシダ類の種分化の過程・機構の解明である。ホウビシダ類におよそどのような種があり、それらがどのような系統関係にあるか、そして各種がDNAの塩基配列レベルでどのくらい違っているかということは明らかにしてきた。したがってどの種とどの種が一番近縁で、しかもどのくらい遺伝的に離れているかもわかっているのである。たとえばヤクシマホウビシダモドキでは、形態学的にもほとんど区別できないものの、rbcLの塩基配列が5つ異なる2つの近縁種が認識できている。これらの種は中国では同所的に存在しているので生殖的隔離はあるのか、もしあるなら、どのような機構で生殖的隔離が起きているかを明らかにできれば興味深いであろう。これを通じて、種分化の研究に発展させられればと考えている。
(村上哲明)