





Kikaithyris hanzawai (Yabe)
腕足動物門テレブラチュラ目
鹿児島県喜界町上嘉鉄
速水格ほか
総合研究資料館、地史古生物部門
これらの「分子化石」は、例えばアミノ酸の光学不活性化(ラセミ化)の度合を用いた年代測定や、窒素の安定同位体を用いた食物連鎖の推定、高分子の構造の比較による古生物の類縁関係の推定など多種多様の研究に応用されており、過去の生物とそのまわりの環境、そしてそれらの時間変遷についての情報の宝庫である。ここでは主に、情報高分子として核酸と並び称されるタンパク質を含む有機物について、その化石からの検出法と生物系統学への応用について簡単に紹介したい。
その前に、研究材料とした腕足動物についての概略を述べる。腕足動物は、背腹2枚の殻を持つ海棲無脊椎動物の一門である(註1)。顕生代を通じて化石として最も普通に見られる生物の1つで、現生種も世界中から知られ、その多くは大陸棚などの比較的深い海底に固着して生息する。化石記録が豊富なこと、現生種の情報も得られること、体サイズも十分大きいこと、貝殻構造が比較的単純で扱いやすいことなどから化石タンパク質の研究材料として適している。もちろん以下で述べる手法は、硬組織の化石を残すその他のすべての生物−化石の古さとしては確実なところで100万年前ぐらいのものまで−に応用可能である。
以下で紹介するのは、喜界島(註2)の湾層(約8万年前)と沖縄本島(註3)の琉球層群(約100万年前)から採集されたキカイチリスの標本に基づいて行った研究の結果である。挿図1に湾層産の標本の一例を示す。キカイチリスは有関節綱テレブラチュラ目ラクエウス科に属する腕足動物で、茎孔(註4)のサイズや殻体の厚さの相対成長の様式(挿図2、3)から、この目の腕足動物としては珍しく成長に伴って固着生活から自由生活へと二次的に生活様式を変化させたことが推察された(註5)。本種に注目したのは、そのような形態的な特異性もさることながら、8万年前の記録を最後にその後は現在の海にいたるまでその存在が確認されていなかったこと、すなわち絶滅してしまったのではないかと考えられたことによる(註6)。絶滅種では通常系統学的な情報源は形態的な特徴に限られるため、絶滅種から分子レベルの情報が得られることの意義は大きい。
 | 21-1 タカハシホタテ化石の産地。研究に使用したサンプルは、Tk:滝川市の空知川河床、In:石狩沼田町幌新太刀別川河床、Sd:仙台市内澱橋付近の各地で採集された。 |
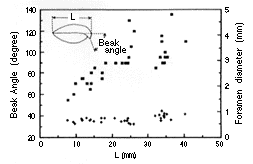 | 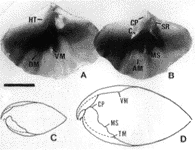 |
| 22-2 Kikaithyris hanzawai の腹殻殻頂部の傾き(Beakangle:黒四角)と、茎孔の直径(Foramendiameter:十字)の腹殻の長さ(L)に対する相対成長。サンプル番号UMUTCB18196。個体数=33。 | 22-3 成体腹殻(A)と背殻(B)の内部および固着生活(C)と自由生活(D)の成長段階を示す個体の縦断面。サンプル番号UMUTCB18196。スケールバーは1cm。各部の名称は省略。自由生活を行う成体では、殻後部の内側(CP, SR, C, HT各部)が肥厚し、殻体がおきあがりこぼしのように底におもりをつけた状態になる。 |
研究対象としたのは、これらの化石標本および比較のために用いた現生の腕足動物40数種の殻(内層と呼ばれる方解石の繊維状結晶から成る部分)に含まれる「結晶内タンパク質」である。骨・歯・貝殻などの生物の硬組織(生鉱物)は、炭酸塩・リン酸塩などの鉱物相とタンパク質・糖質などの有機物から成り、これらの有機物は生鉱物の形成に積極的な役割を果たしていることが知られる。これらの有機物は、鉱物結晶との位置関係から、(1) 結晶間にマトリックスとして存在するもの、(2) 結晶表面に強く結びついて存在するもの、(3) 結晶内に存在するもの(結晶内タンパク質)に分けることができる。生きた硬組織中の存在量は、通常(1) が圧倒的に多い。しかし、結晶と結びつくことにより化学的な安定性が増すことや、まわりを結晶に囲まれることにより水分・バクテリアなどに対する物理的な防御が得られることから、(2) や (3) の方がむしろ化石に残る可能性が高いことが予想され、また実際それを裏付ける例も知られるようになってきた(註7)。
内層に含まれる結晶内タンパク質を抽出するにはまず、殻表面に付着している堆積物や表生生物などの異物を歯ブラシやメスなどで除去したのち水洗し、10パーセント程度の濃度のブリーチ(次亜塩素酸ナトリウム水溶液)に漬け2日ほど室温においておく(註8)。軽く蒸留水で洗ったのち、標本をプラスチック製の遠心チューブ内で適当な棒を用いて完全につぶし、ブリーチを微量に含む蒸留水を加えてこなごなになった殻断片を懸濁させる。ここで緻密な結晶からなる外層の破片は先に沈澱してしまうので、上澄みを静かに別のチューブに移すことにより懸濁している繊維状の内層結晶を分離する。遠心分離により内層結晶片を回収し、蒸留水(註9)で沈澱を洗う、という作業を10回ほど繰り返すことによりブリーチを完全に除去する。最終的に、遠心分離により繊維状結晶を回収し、凍結乾燥したのち(註10)、一定量の結晶粉末を一定量のEDTA水溶液によって脱灰することにより、結晶内タンパク質を溶出させる(註11)。
このようにして抽出した化石抽出物から目的とするタンパク質を検出し、その構造を近縁現生種のものと比較する方法として免疫学的方法を用いた。免疫学的方法は、タンパク質の小さな領域(抗原決定基)を特異的に認識する抗体を利用して、目的とするタンパク質(抗原)を検出する方法である。この方法の利点として、もとのタンパク質が多少分解していても検出できる点や、感度の高い免疫測定法(註12)を利用することにより少量の化石タンパク質を検出できる点、抗原−抗体反応の特異性から、バクテリア等に由来する汚染物を区別できる点などをあげることができる。
免疫学的方法を用いるためには、まず目的とするタンパク質に対する抗体を用意しなければならない。そのためには、先ほどの要領で抽出した抽出物を脱塩し、タンパク質を精製したのち、適当な実験動物に対して免疫注射を繰り返す必要がある(約2ヶ月ほどかかる)。本研究ではウサギを用いて、キカイチリスとの近縁種を含む、現生15種の腕足動物の殻から抽出した結晶内タンパク質に対する抗体をそれぞれ作成した。十分な抗体価をもった抗体の作成には、先ほど述べた繊維状結晶粉末の重量にして約20グラムの殻が必要である(註13)。しかし、このようにして得た抗体を用いれば、現生標本の場合、数ミリグラム(耳かき一杯ほど)の結晶粉末から、すなわち一個体のわずかな破片からでも、目的とするタンパク質の検出が可能である。後に述べるキカイチリスの化石標本を用いたアッセイにおいても、最終的に結果を得るために必要であった殻の量は標本一個体分に満たない。
さて、抗原−抗体反応の強さの違いから、抗原タンパク質の全体的な構造的類似性を分類群間で比較し、その類似性に基づいて系統関係を推定する方法が免疫学的方法であるが、この方法を化石に応用する前に、現生種において系統学的に意味のある結果が得られるかどうかをまず確かめた。挿図4にテレブラチュラ目のすべての超科と大部分の科を代表する30属の現生腕足動物について行った結晶内分子の免疫学的比較の結果を示す。また挿図5は、さらに詳細な実験を行った結果を概念的にまとめたものである。4つの主要なグループが認識されたが、これらのうち2つ(TuとC)は、従来の形態に基づく分類体系での超科にまさに対応していた。残る2つは従来の分類体系では同一の超科にされていたが、形態的に一方は典型的なもの (Te)、もう一方は特異な形態をもつもの (K) として一応の区別がなされていたものである。この特異な形態をもつグループ (K) が、明らかにTeではなく、Tuグループに近いという結果は驚きであったが、形態学的にも解釈は可能であり、従来の系統の解釈の再検討をうながすものであった(註14)。
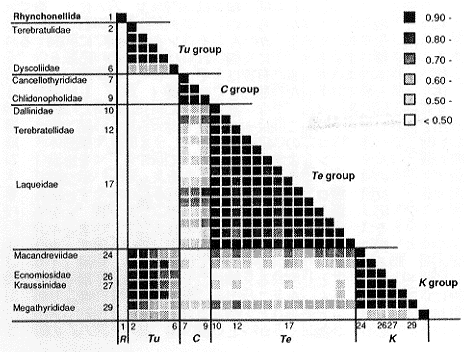 | 22-4 免疫学的データに基づく現生腕足動物30種間の類似度。値が大きいほど類似性が高いことを示す。縦軸横軸の数字は各種に付けられた番号。4つの主要なグループ(Tu, C, Te, K)がテレブラチュラ目内に認識され、TuグループとKグループの間に高い類似性があることが示された。 |
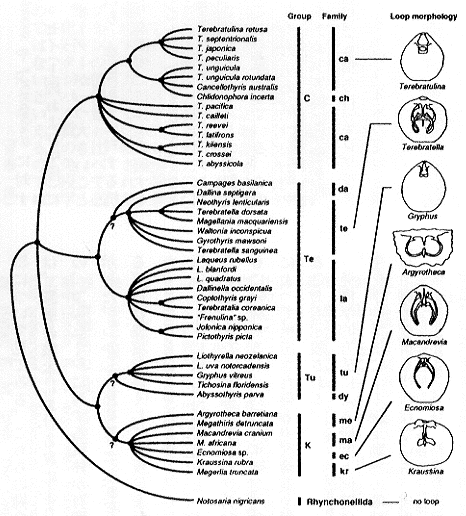 |
22-5 免疫学的データからみた現生テレブラチュラ目腕足動物の類似関係。代表的な属の腕骨(形態学的分類において重視される石灰質の構造物)のスケッチ(Loopmorphology)を横に示す。縦軸横軸の長さはともに類似度とは比例しない。腕骨の相対的長さで2つの亜目に分ける従来の分類体系は支持されなかった。 |
現生種において、結晶内タンパク質の免疫学的比較が系統学的に有効であることは明らかとなったが、この方法を化石に応用することは必ずしも容易ではなかった。現生種を用いたアッセイでは、それぞれの一次抗体(腕足動物の結晶内タンパク質に対する抗体)は、それぞれの抗体において最適な一定の濃度に希釈して使用した。同様の方法で化石抽出物に対するアッセイを行っても、有意なシグナルを得ることができなかったのである。抗原として用いる化石抽出物の量を増やしたり、一次抗体の濃度を増やしたり試行錯誤を繰り返したのちに開発した方法が、連続抗体濃縮法 (SerialAntibodyConcentrationMethod: SACM) である(註15)。
この方法は、一定量の抗原(化石抽出物)に対して、一定間隔で低濃度から高濃度まで異なる濃度(通常10種類の異なる濃度)に希釈した一次抗体を用いた一組のアッセイを行う方法である。ある抗体に対する化石抽出物の免疫反応の強さは、化石抽出物とのアッセイにおいて、その抗体を準備した現生種からの抽出物とのアッセイと同レベルの免疫反応を得るために、どれだけ(何倍)その抗体を濃縮する必要があったか、その濃縮率で表すことができる。
挿図6に連続抗体濃縮法をキカイチリスの化石に応用した結果を示す。まず湾層産約8万年前の化石 (Fossil 1) について。Laqueus、Pictothyris、Waltonia 各属の現生種の結晶内タンパク質に対してそれぞれ作成した抗体は、それぞれ濃度を高めることによって(横軸の左側ほど高濃度)、化石からの抽出物と明らかに有意な反応を示すようになることがわかる。また、このように抗体の濃度を高くしてもネガティヴコントロールはほとんど無反応のままである。
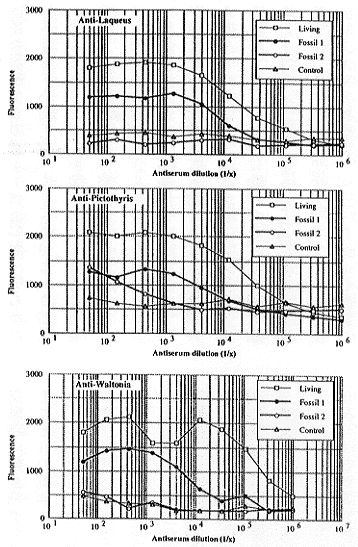 | 22-6 Kikaithyrishanzawai 化石(約8万年前と約100万年前)からの免疫学的反応曲線。 Kikaithyris と同じラクエウス科に属する現生 Laqueusrubellus(上段)と現生 Pictothyrispicta(中段)、およびテレブラチュラ科に属する現生 Waltoniainconspicua(下段)に対する抗体を用いて、連続抗体濃縮法により化石タンパク質を検出した。横軸:抗体の希釈度。縦軸:蛍光の強度(免疫測定法によって最終的に測定される免疫反応の強度)。Living:それぞれの抗体を作成した現生標本。 Fossil1:湾層産 Kikaithyrishanzawai(約8万年前)。Fossil2:琉球層群産 Kikaithyrishanzawai(約100万年前)。Control:湾層産二枚貝。それぞれのプロットは2つの独立した測定の平均値を示す。 |
現生標本と同レベルの反応を得るために必要な抗体の濃縮率を見てみると、Waltoniaに対する抗体を用いたアッセイ(下段)における濃縮率は、LaqueusやPictothyrisに対する抗体を用いたアッセイ(上段および中段)における濃縮率よりオーダーが1つ大きいことがわかる。すなわち、Waltoniaに対する抗体を用いた方がより高濃度に抗体を濃縮する必要があり、このことはLaqueusやPictothyrisの方が、Waltoniaよりも結晶内タンパク質の構造においてキカイチリスとの類似性がより高いということを意味している。このことはまた、キカイチリスがLaqueusやPictothyrisと同じラクエウス科に分類され、Waltonaが同じ超科でも別の科(テレブラチュラ科)に分類されるという従来の見解と調和的である。
次に琉球層群産約100万年前の化石 (Fossil2) について見てみる。こちらの場合においては、湾層の標本における反応より弱いものの、Pictothyrisに対する抗体が有意な反応を示した。しかしLaqueusとWaltoniaに対する抗体は濃度を高くしても反応を示さなかった。このことは、キカイチリスが形態的にはPictothyrisに最も近縁であるという考え(註16)と符合する。また、結晶内タンパク質は時間とともに分解するが、約100万年経たのちも、抗体によって検出され得る一次構造を保持していることも示唆される。
免疫学的方法による研究と同時に、様々な生化学的手法(註17)による腕足動物の殻結晶内タンパク質の分析も行われている(註18)。現生のNeothyrislenticularis には、6.5キロダルトン (kDa)(註19)、16キロダルトン、47キロダルトンの3つの主要なタンパク質が、また、Terebratulinaretusaには、30キロダルトンのタンパク質一種類だけが含まれることが確かめられている 。6.5キロダルトンと47キロダルトンのタンパク質については、N末端からそれぞれ30と10残基のアミノ酸配列も決定された。いずれもこれまでに知られていない配列である。しかしこれらのタンパク質の全構造と機能の解明については、これからの研究に負うところが大きい。
化石タンパク質は、過去の絶滅生物の系統関係の推定において有用である。また、生体分子がどのようなプロセスを経て進化したのか、またその過程でどのような制約が働いたのかなどについて、他から得られない知見を提供するかもしれないという可能性を秘めている。しかし残念ながら、化石から最初にアミノ酸が報告されて40年あまり、DNAが報告されて10年あまりが過ぎるが、現状では、分子進化に関するこの種の有益な知見はまだ得られていないといってよい。
免疫学的データは、少なくとも過去100万年の分子進化の過程を、腕足動物の化石を用いて連続的にたどることができる可能性を示す。しかし、そのためには、化石タンパク質の一次構造の情報を得ることが必要条件であると言える。これは構造決定に必要な純度の化石タンパク質を精製することが困難なため必ずしも容易ではないが、アミノ酸の側鎖が保存され抗原決定基を保持したペプチドが存在するならば、その一次構造を決定することは原理的には可能であろう。1990年には、3600年前のモアの化石骨に含まれるタンパク質の一次構造が一部決定されている(註20)。このような研究がルーチン化されるようになれば、化石タンパク質による分子進化の研究も新しい局面を迎えることになろう。
最後に、最近世間を賑わせている化石DNAについて、化石タンパク質の免疫学的研究との関連において簡単にコメントしておく。遺伝子の本体であるDNAが化石から抽出復元されることの意義は大きい。もし仮に、どのような化石からも任意のDNAを容易に増幅できるとすれば、遺伝子の2次産物であるタンパク質の化石の研究は、少なくとも系統学的応用という目的においてほとんどその意味を失ってしまう。 しかし現実には、そもそもDNAが残り得る化石は、タンパク質が残り得る化石の集合のごく一部を占めるに過ぎない(註21)。このことは化石の多くが外骨格という、細胞を含まない(しかしタンパク質は含む)、いわば「死んだ」組織で占められることにもよるが、また同時にDNAよりもタンパク質の方が、一次構造をつくる化学結合やポリマーを構成する要素の安定性が高いことにもよる。現在までのところ、おそらくこの世において万人が認める最古のDNA(ポリヌクレオチド)は数万年、タンパク質(ペプチド)は100万年前後の古さと見てよいのではないかと思う(註22)。
また、確実にDNAが残っていると思われる化石においても、増幅が可能なDNAはミトコンドリアDNAなど、1つの細胞あたり多数のコピーが存在するものに限られ、また1回に増幅できるDNA断片の長さも200塩基対程度である。これは系統学的な情報量としては、例えば免疫学的方法で比較されたコラーゲンやアルブミン(註23)の情報量と比べて必ずしも大きくない。化石タンパク質の免疫学的研究も捨てたものではないといえるのではないだろか。
(遠藤一佳)
註1 腕状の摂食・呼吸器官が、外観の類似する二枚貝の足と相同であると最初に誤認されたためこの名がある。