





Fortipecten takahashi (Yokoyama)
軟体動物門二枚貝綱翼形亜綱ホタテガイ科
北海道空知支庁滝川市
岩崎泰頴(UMUT CM 18117a)
総合研究資料館、地史古生物部門
しかし、見かけの類似は決して意味のない現象ではない。多くの場合、機能上の共通性があるからこそ見かけの類似が生じるのである。したがって、化石生物の機能形態学では見かけの類似を逆に利用して研究を進めるのが普通である。生活様式がよく分かっている現生生物と体や器官の形を比較することにより、化石生物の形態の機能的意味をある程度推定することができ、古生態を復元することが可能となる。例えば、恐竜や化石哺乳類の食性は歯の形からかなり詳しく知ることができる。類似の相手は人工物であってもかまわない。種々の人工物は最も効率が良くなるように造られるから、生物の体形や器官の構造が人工の道具や機械に類似していることは、機能を推定するよい手がかりとなる。
ところで、白亜紀以前の平坦な海底は、貝類にとってまだ強力な捕食者が少ない平和な環境であったらしい。これは現在の海にはほとんど見られない横臥型(平坦底にただ寝そべって生活するタイプの)二枚貝類が豊富であったこと、殻口部が肥厚したり武装的な彫刻をもつ巻貝類が少なかった事実によっても暗示されている。貝類に対する捕食圧が高まった原因としては、貝類の殻を破壊して食べる硬骨魚類や十脚類、二枚貝の殻をこじあけて食べるヒトデ類、殻に穴をあけて食べる巻貝(特にタマガイ類)が白亜紀中頃以降に急増したことがあげられている。
G・J・ベルメージという研究者は、この白亜紀ごろに起こった浅海における捕食圧の急激な増大を「中生代の海洋変革」と名づけた。さらに、捕食者と被食者が互いに攻撃と防御の能力を高めあうことを「エスカレーション」と呼んだ。この一連の学説は、さらに検証を要する点があるが、広く一般に受け入れられ、貝類の形態進化の重要な原因になったと考えられている。実際にこの学説は、古生物研究者が長年にわたって積み重ねられてきた広汎な進化現象(特に方向性のある貝類の形態i化)をよく説明するものである。ホタテガイ類の遊泳行動も1つの有効な対捕食者戦略であり、この仲間は遊泳能力を獲得したことによって、通常の二枚貝類にとって極めて危険な平坦底の表層でも生活することが許されたと考えられる(挿図1)。
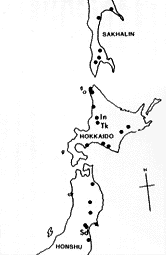 | 21-1 タカハシホタテ化石の産地。研究に使用したサンプルは、Tk:滝川市の空知川河床、In:石狩沼田町幌新太刀別川河床、Sd:仙台市内澱橋付近の各地で採集された。 |
筆者らがタカハシホタテに特別の関心を抱いたのは、この種が新生代には珍しい横臥型の生活者ではなかろうかと考えたからである。タカハシホタテの幼貝は形態的に現生のホタテガイの幼貝と大きな違いはないように見える。多分ホタテガイと同様に遊泳したと思われる。しかし、成貝の形状はホタテガイと著しく異なる。横山博士もサハリンの同じ産地から出た小型の個体(おそらく幼貝)をPecten agnatusと命名し、当初は別種と考えていた。しかし、成長の中期(殻径7センチぐらい)に達すると、成長のパターンが急に大きく変化する。成貝の右殻は半球状に強く膨れ、殻質は左右ともほぼ同じサイズのホタテガイに比べ4倍以上も厚くなる。
タカハシホタテの成貝は非常に重厚なので遊泳できたとは到底思えない。別に行ったホタテガイ類の流体力学実験の結果を外挿すると、殻の重量1キログラム(水中の重量は約600グラムになる)、殻径16センチのタカハシホタテの個体が水中重力を越える揚力を得て遊泳するには、最適の迎え角をもって泳いだとしても、秒速1.2メートル以上のスピードを必要とする。ちなみにこれまで観測された遊泳速度の最大はナンヨウツキヒの秒速1.6メートルであるが、一般の遊泳種の速度はこれよりもずっと遅く、ホタテガイではおそらく0.6メートル以下である。
タカハシホタテは絶滅種であるから、その生態を直接観察することはできない。このような場合、相対成長のパターンを生態のよく分かった種と比較して、生活様式を間接的に推定するのが古生物研究者の取り得る道である。ここでは成長様式をホタテガイと詳しく比較することによって、「横臥型」の仮説を検証することにした(挿図2、3)。
 |  |
| 21-2 タカハシホタテの成熟固体(4方向から撮影 ×0.65)。産地:滝川市の空知川河床(岩崎泰頴採集)。標本番号UMUTCM18117a。 | 21-3 タカハシホタテの幼期個体(3方向から撮影 ×1)と中期個体(3方向から撮影 ×0.65)。産地:石狩沼田町幌新太刀別川河床(速水格採集)。標本番号UMUTCM18116a, b。 |
一般に遊泳型のホタテガイ類では、成長に伴って多少とも殻や軟体部のプロポーションが変化する。しばしば見られる傾向として、
(1) 殻を相対的に薄くして、水中重力を軽減する。
(2) 殻の前背縁・後背縁の隙間を広げて大量の水を噴出する。
(3) 末広がりに殻頂角を大きくしたり、耳状部を相対的に小さくして水流噴出の効率を高める(足糸付着型の種のように殻頂角が小さいと、前後の水流の力が打消しあって効率が悪いと考えられる)。
(4) 前後に広い殻に成長して揚力/抗力の比を大きくする(飛行機の翼と同様にアスペクト比を大きくする効果がある)。
(5) 閉殻筋の速筋(横紋筋)の部分を相対的に大きくして、より強力な水流を噴出できるようにする。
(6) 閉殻筋が内表に対して斜めになり、より急速な閉殻を可能にする。
などがあげられる。これははいずれも個体が成長しても遊泳能力を維持する上に有利な調整であると考えられる。ホタテガイの成長にも確かにこのような傾向が認められる。
成長に伴って増加する生物の2つの計測部位 (x, y) はべき関数 y=bxa によくあてはまることが知られている。これが相対成長の一般式(アロメトリー式)である。この一般式はlogy=alogx+logb と同等であるから、多数の個体からなるサンプルについて平均的なアロメトリー式を求めるには、それぞれの計測値を対数変換した上で直線回帰(生物測定学では通常の最小2乗法ではなく、reduced major axis 法がよく用いられる)を行えばよい。この式にしたがうあらゆる成長パターンは、両対数グラフ上では直線で表現され、aの値は直線の勾配で表される。なお、a=1 は等成長(プロポーションの変らない成長)を意味し、a≠1は不等成長(狭義のアロメトリー)であることを示す。ただし、xに長さ、yに体積や重量をとった場合には、a=3が等成長になる。
相対成長を解析した結果、タカハシホタテの成長パターンはホタテガイや他の遊泳性のホタテガイ類と著しく異なることが明らかとなった。その特徴を前述の6項目の一般的傾向と対照して示すと、次のようになる。
(1) 殻高と殻重量の関係 幼期(殻径70ミリ以下)はほとんど等成長 (a≒3) でホタテガイとほとんど違わないが、中期以後はホタテガイが殻をやや軽くする方向に変化するのに対して、タカハシホタテは著しく殻を重くする方向に成長パターンが急激に変化する(挿図4)。
(2) 前背縁・後背縁の隙間 タカハシホタテにも幼期には隙間があるが、中期以後はほとんど閉じる(遊泳しないホタテガイ類では隙間の存在は他の生物に入り込まれるので不利であろう)。
(3) ホタテガイでは成長に伴って殻頂角が大きくなるのに対して、タカハシホタテではむしろ小さくなる(挿図5)。また、耳状部の相対的な大きさは、ホタテガイではほとんど変らないのに対して、タカハシホタテでは著しく大きくなる(挿図6)。
(4) ホタテガイでは成長に伴って殻長/殻高の比(アスペクト比)がやや大きくなるが、タカハシホタテではむしろ小さくなり、殻は縦長になる。
(5) ホタテガイでは成長に伴って閉殻筋の速筋の部分が相対的に大きくなるが、タカハシホタテでは逆に遅筋の部分が大きくなる。
(6) ホタテガイの閉殻筋(速筋)は殻の内面に対してかなり斜めに付着するが、タカハシホタテの成貝ではほとんど垂直に付着する(挿図7)。
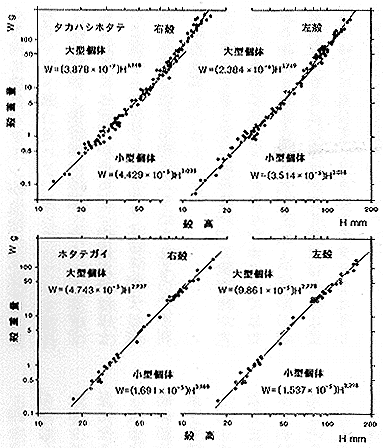 | 21-4 タカハシホタテとホタテガイの殻高(H)と殻重量(W)の関係。タカハシホタテの相対成長は明らかに2相アロメトリーを示し、成長の中期以後に殻が急速に重厚になることが分かる。 |
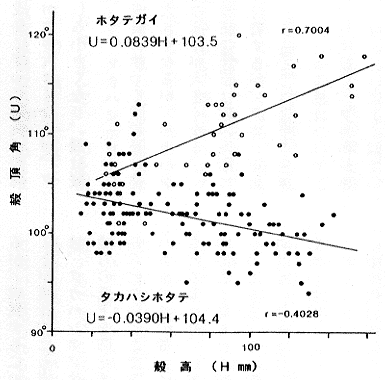 | 21-5 タカハシホタテとホタテガイの殻高(H)と殻頂角(U)の関係。ホタテガイは成長に伴って殻頂角が大きくなるが、タカハシホタテは逆に小さくなる。相関はあまり高くないが、いずれも有意である。 |
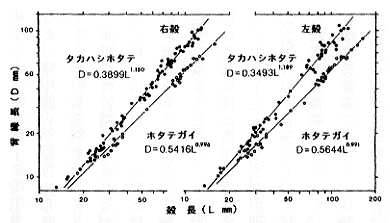 | 21-6 タカハシホタテとホタテガイの殻長(L)と背縁長(D)の関係。背縁長は耳状部の大きさを表す。幼期にはあまり差がないが成長するにつれてタカハシホタテの耳状部が非常に大きくなることが分かる。 |
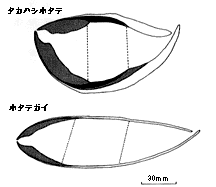 | 21-7 タカハシホタテとホタテガイの殻の垂直断面。タカハシホタテの殻の外層(白地の部分は成長の中期以降に急速に厚くなる)。ホタテガイの閉殻筋(点線部)は殻表に対してかなり斜めにつくが、タカハシホタテではほぼ垂直である。 |
以上の成長パターンの比較で、タカハシホタテとホタテガイの幼貝は互いによく似ているが、成長するにつれて形態差が大きくなることが統計的にも立証された。注目されるのは、タカハシホタテの成長の中期以降に遊泳能力の低下を防止するような調整がまったく見られないことである。それどころか、タカハシホタテの殻および閉殻筋の成長過程に見られる変化は、いずれも遊泳型のホタテガイ類に広く見られる傾向とはまったく相反するものである。特に殻重量の急激な増加は、中期以後に遊泳能力が急速に失われたことを示している。
「氷山戦略者」がしばしば半球状の殻形をとるのはなぜだろうか。1つの説明は、このような椀状の形態をとることにより、呼吸や採餌に必要な開口部(二枚貝では左右の殻の接合面)を底質に埋もれることがないように高い位置に保つことができる利点である。また、動物体の重心が半球状の殻の内部にあれば、水流や他の動物によって生息姿勢を乱されても、「起きあがりこぶし」のように重力だけで姿勢を復元できるメリットがあったと思われる。
中生代の二枚貝には「氷山戦略者」が多いと考えられている。しかもこれは系統とは無関係に様々な目や科に出現している。最も有名なのは、進化研究の好材料となっている自由生活型のカキ類で、ジュラ紀のGryphaea、白亜紀のExogyraやPycnodonteの一部の種である。いずれも成貝は自由生活し、半球状に強く膨れた重厚な左殻を下側にして柔らかい泥底に「浮かんで」いたらしい。イノセラムス類のVolviceramusなどについても同様の生活様式が考えられている。
ホタテガイ類では、ジュラ紀のWeyla、白亜紀のNeitheaなどが「氷山戦略者」であったと考えられる。これらはいずれも半球状の右殻とほとんど偏平な左殻を有し、遊泳型のホタテガイ類に比べて、殻は重厚でやや縦長のことが多い。また、足糸湾入部は退化しているので、自由生活者であったことも疑いない。興味深いのは、タカハシホタテの殻の形状がこれらの中生代のホタテガイ類に極めてよく類似していることである。もちろん、これは時期を違えて起こった並行進化による見かけの類似と考えるべきだろう。Neitheaは白亜紀末に(太平洋海域では白亜紀中頃に)子孫を残すことなく絶滅しているので、鮮新世のタカハシホタテとの間に直接的な系統関係があるとは考えられない。
先に触れたように、ホタテガイ類の遊泳速度が殻のサイズとは独立であると仮定すると、殻の表面積(単位cm2)を殻重量(単位g)で除した値は形態から遊泳能力を推し量る1つの目やすとなるであろう。遊泳時の揚力は表面積に比例し、動物体の水中重力は殻重量の六割程度と見積ることができるからである。計算を簡単にするために、殻の表面積を殻長×殻高で表して、いくつかの種についてこの比を求めてみた。もちろん、どの種でもこの比は成長に伴って次第に小さくなる。最も活発に遊泳するツキヒガイでは、老成個体でもこの値が1.8以上である。遊泳性のホタテガイやイタヤガイではこれよりも小さいが、老成しても1.0を下まわることはほとんどない。これに対して、タカハシホタテは成長の中期(殻径7〜8センチ)で1.0を割り、老成すると0.3〜0.4にまで低下する。種によって遊泳速度は異なるのであくまで目やすに過ぎないが、1.0程度が遊泳ができる限界であろう。また、この値が大きくても、揚力に対して抗力が大きいと活発な遊泳は困難になるだろう。流線形のツキヒガイが最適の迎え角で水平に遊泳すると、揚力/抗力の比が5.0以上になる。タカハシホタテの成貝のようなごつごつした物体を水中で動かすと、流線の剥離や渦を生じ、大きな抗力が生じると予測される。
タカハシホタテは、幼期には現生のホタテガイと同様に遊泳によって捕食を避けていたが、成長の中期(殻径約7センチ)に達すると、突然この戦略を放棄して「氷山戦略」に切り替えた。いわば、捕食者に対して殻を重厚にして、「食えるものなら食ってみよ」と居直ったのである。この戦略は一時的には成功して、本種が北西太平洋の冷水域(東北日本、サハリン、カムチャッカ)に分布を広げた時期があった。これは中生代型の「氷山戦略」の復活と見ることができよう。捕食圧が相対的に低いと見られる冷水域でこの戦略が復活したのは興味深い。ただし、足糸付着型から直接横臥型になったのではなく、遊泳型を経て二次的に横臥型に変身した点では中生代の類似種と異なる。
タカハシホタテの祖先は、中新世に広く繁栄していた遊泳型のホタテガイ類(おそらく広義のPatinopecten属)に求められるだろう。また、遊泳型から横臥型への生態の変化は瞬時に起こったと考える必要はない。この変化は、最初に遊泳型のある種の成長末期に現れ、次第に変化が起こる時期が成長中期に繰り上がったのではないかと思う。こう考えるのはいくらか理由がある。実はタカハシホタテにはかなり著しい地理的変異がある。時代の前後関係は明らかではないが、仙台付近の竜の口層や北海道太平洋岸に産するタカハシホタテの個体は、北海道中部やサハリン産の個体に比べると、右殻の膨らみが弱く、殻が急速に重厚になる時期は殻径が約10センチに達してからである。つまり、半球状の典型的なタカハシホタテに至る中間の段階を表していると考えることができよう。
(速水 格)
小文は下記の研究論文の概要に若干の解説を加えたものである。用いた標本はすべて東京大学総合研究資料館地史古生物部門に登録保存されている。