





Calyptogena (Calyptogena) soyoae Okutani
軟体動物門斧足綱オトヒメハマグリ科
相模湾750〜1400m
1987年採集
太田秀(しんかい2000にて採集)
海洋研究所海洋生物生態部門
深海の生態系にこの図式を当てはめると、はるか数1000メートル上層の有光層で植物プランクトンたちによって、あるいは数100キロメートル離れた陸上の植物によって作られた有機化合物が、長い水柱の旅を経て海底に到達し、そこで待ちかまえていた底生生物の食物となって無機物に分解される。表層から海底に達するまでの間にかなりの部分が分解・消費されてしまうため、深海まで到達する有機化合物の量はたいへん少なく、したがって深海生態系というものは生物の極めて希薄な世界であると思われていた(海底に達する有機化合物の量を直接測定しようという試みが、セディメントトラップと呼ばれる測器を用いて行われ、表層で生産された有機化合物のせいぜい1パーセント程度しか海底には到達しないという結果が出ている)。
このような深海生物学の常識を背景に、1970年代にガラパゴス諸島沖の水深2600メートルの海底で、アンガスと呼ばれる頑丈なカメラシステムや、深海潜水艇アルビンを用いて大洋底拡大軸を直接観察しようとしたアメリカの地球科学者たちがみたものは、想像を絶するものであった。そこには巨大な二枚貝類と、白い管から真っ赤な触手を出している得体の知れない生物とが異様な高密度で密集しており、その生物量は一般の深海底よりも4桁程高いものであった。この高い生物量を支えているものが、光合成一次生産者ではなく、化学合成一次生産者であることが証明されるのに、長くは要しなかった。
その後、世界各地の大洋底拡大軸、マリアナなどの背弧海盆およびマイクロプレート型のリフト系などで、続々と同様な熱水生物群集が見出された (Desbruyeres et al.,1994)。また、熱い温泉水ではなく冷たい湧水に支えられた類似の生物群集がオレゴン沖に発見され (Suess et al.,1994)、ほとんど同時にわが国周辺でも初島沖、日本海溝、南海トラフなどのような海洋プレートの沈み込み帯の陸側斜面で、しんかい2000やノチールなどの有人潜水調査艇で発見された。さらにプレートテクトニクスとは特に関連せずに、地下からなんらかの物質が湧出する条件が整っている海域にも、独立栄養細菌に依存する生態系が見出された。例えばルイジアナ沖の天然ガス浸出地帯(Childress et al.,1986)、グランドバンクスに見られる海底地すべり層から重力的に絞り出される冷湧水海域(Mayer et al.,1988)などである。このように、現在では世界各地に地学的エネルギーに依存した生態系が発見されており、シロウリガイやハオリムシ、シンカイヒバリガイなどがこの種の生態系の代表的な動物群であることが判明している(挿図1)。ここでは熱水噴出孔生態系のシロウリガイ類を中心に据え、その自然史に加えて、様々な化学的独立栄養過程をエネルギーの根幹とする生態系について解説する。
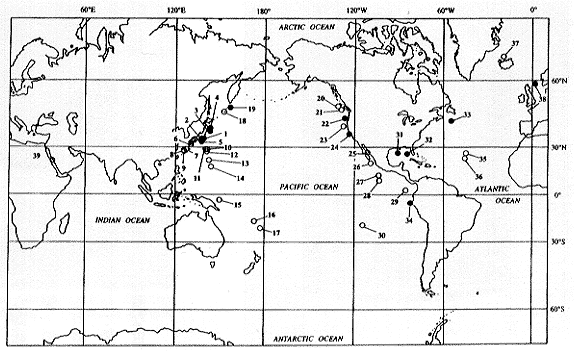 | 20-1 世界の熱水N水生態系の分布 |
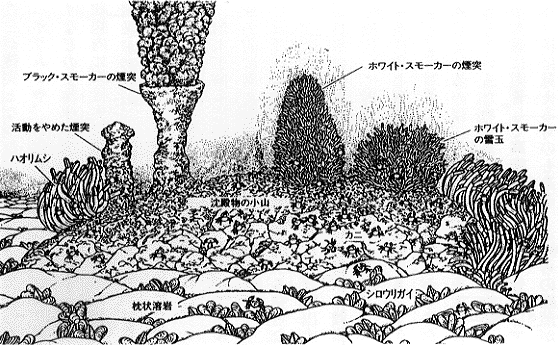
20-2 熱水生態系の概念化した光景。高温のブラックスモーカーからは350℃に達する硫化物に富んだ黒い熱水が吹き出していて、煙突それ自身も硫化物の沈澱物から出来ている。ホワイトスモーカーから放出される熱水は300℃以下である。チムニーのまわりは、イトエラゴカイ(Paralvinella)属の多毛類が密集している。またハオリムシもその周辺で集群を作り、裾野にはユノハナガニ類が群れる。また、海底下で海水と混合して散開的に生ぬるい温水としてわき出す水は自由生活をしている独立栄養細菌を含んで白く白濁している。やや離れた枕状溶岩の割れ目には、シロウリガイが足を差し込んで多数生息している。この深海のオアシスは熱水の影響のない周辺の海域の1万倍以上の生物量を持ち、そのエネルギー源は熱水から供給される硫化水素を酸化的に利用する独立栄養細菌の生産する有機化合物である(図は日本語版『サイエンス』1981年7月号より)。
熱水生態系が発見された当初、砂漠のように生物の希薄な深海の環境に突如現れるこの深海のオアシスが、いかなるエネルギー源によって支えられるかに注目が集まった。2つの仮説が提出された。一つは独立栄養細菌説であり、もう一つは熱水の起こす熱対流によって、有光層や陸で光合成で生産された有機化合物が収束するというものであった。あの巨大なハオリムシに口も消化管も存在しないことから考えても、化学合成一次生産者の共生を仮定する考え方の方に分があることは明らかだった。これだけの現存量を支えるエネルギー供給源は海底下に求めざるを得ない。
独立栄養細菌説を支持する根拠が続々と報告された。まず、乳濁した海水に酸化可能な硫黄イオンを添加すると一次生産力が有意に増加した。また、独立栄養細菌の代謝経路にのみ存在する酵素が確認された。一方、底生生物自身については、生物の軟体部の13C:12C比や15N:14N比が独立栄養細菌の有機物に特徴的な値に近いことが明らかとなり、細菌が底生生物を作り上げる材料となっていることが確実となった。消化管を全く欠如したハオリムシや、消化管が退縮したシロウリガイなどの生物体を電子顕微鏡で観察すると、細胞内に原核生物像が充満し、細菌との共生関係が形態的に証明され (Cavanaugh, 1983, 1985)、酵素的、分子遺伝学的 (Kim et al., 1995) にも化学合成細菌との共生が証明された。
熱水生態系に豊富に存在し、これをエネルギー的に支える細菌類は、3つのカテゴリーに分けられる。まず、水中に懸濁する完全な独立栄養細菌があげられる。かれらは噴出する熱水に含まれる硫化物を、周辺海水の酸素で酸化して得られるエネルギーを利用して、炭酸同化と有機物生産を行い増殖する (Utsu-mietal., 1994)。散開型の低温噴出水がやや白濁しているのは、この独立栄養細菌を大量に含んでいるためである。また大型の生物体上や海底に固着して高密度に増殖し、分厚いマット状に見えるものもある。これらの細菌が作り出す懸濁態の有機化合物、ならびに細菌体自体がシンカイヒバリガイ、ミョウガガイ類などの濾過食者や、笠貝類のようなグレーザーの重要な食物となっている。つぎに、底生生物の体内に共生し、宿主から硫化水素と酸素と二酸化炭素の供給を受けて増殖し、生産した有機化合物を底生生物に与えている独立栄養細菌がある。シロウリガイでは消化管が退化的であるし、ハオリムシに至っては全く存在しない。前述のように深海の環境は基本的には食物の乏しい環境であり、これらの大型生物が摂食機能を全く持たなくとも高密度に生息できるのは、ひとえに体内の共生細菌のおかげである。最後に、硫化物や炭化水素の酸化エネルギーを利用しているが、それは従属栄養的代謝の補助手段に過ぎず、独立栄養と従属栄養を使い分けているものもいる。
いずれにしても熱水生態系に生息する独立栄養細菌のほとんどは、硫化水素酸化型とメタン酸化型であることがわかっている。つまり深海のオアシスは、清冽な泉水ではなく、高温で黒々とした温泉水に含まれる毒物(硫化水素)そのものを「命の泉」としているのである。
深海のオアシスを支える泉は、新たに海底を作り出す場である大洋底拡大軸に沿って点在している。拡大中軸谷には無数の断層が並行し、その割れ目にそって海水が地中にしみこむ。地中深く到達した海水は熱いマグマ(結局は地球内部でウランなどの放射性壊変によって放出されたエネルギー)と高温高圧の条件で反応する。この時最も重要なのは海水の硫酸塩と岩石中の鉄とが反応して硫化水素と鉄酸化物を生成する過程である。この反応後上昇して海洋底に戻ってくる熱水は硫化物に富み、それら(特に硫化水素)は独立栄養細菌に基質として利用され酸化されて硫酸塩に戻る(挿図3)。つまり放射壊変によって発生する熱エネルギーが地中深くに存在する化学工場で硫化水素という無機物の形の化学的エネルギーに変換され、それを海底で独立栄養細菌が有機化合物に変換し(生産者)、それを食物としてシロウリガイなどの巨大な生物(消費者)が維持されるのである。
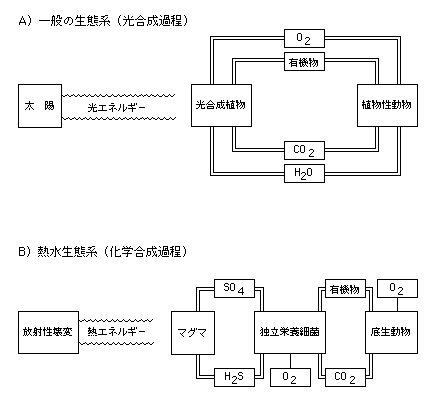 |
20-3 一般の生態系は太陽から来る光エネルギーを利用した光合成過程によって生産された有機物に支えられている(A)。しかし熱水生態系では海底の割れ目からしみこんできた海水がマグマに接し、硫化水素など硫化物を多量に含んで海底に噴出する。この硫化物を酸化して得たエネルギーを利用して独立栄養細菌が生成する有機物(化学合成過程)がすべての生物のエネルギー源となる(B)。 |
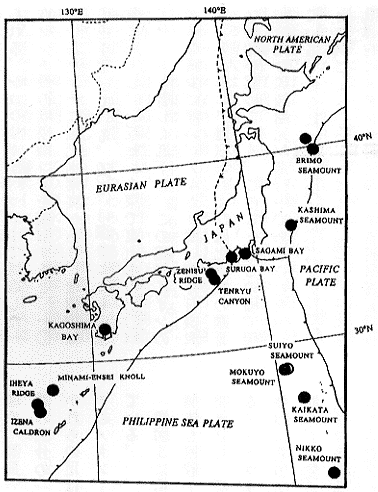 |
20-4 わが国周辺における熱水N水生態系の分布 |
初島沖の冷湧水生態系は、海洋科学技術センターの有人潜水作業艇しんかい2000を用いて、わが国の研究者によって学際的な研究が精力的に進められており、かなりいろいろなことがわかっている。冷湧水生態系と熱水生態系の大きな相違点は、後者が完全な独立栄養の生態系であるのに対し、前者は過去に生産された有機物にそのエネルギー源を依存している点であろう。我々人類が化石燃料に大きく依存して生活しているように、かれらも非常に古い有機物由来の炭素を利用していることが湧出する低分子炭化水素ガス成分比や生物体の14C値や13C:12C比から確実になっている (Saino&Ohta, 1987)。一方、ここに生息するシロウリガイは熱水生態系のものと同様に、硫化水素酸化型の独立栄養細菌を共生させている (遠藤ら1987、Endow&Ohta, 1990)。
挿図5に示すように、シロウリガイはどの個体も粗砂〜細礫サイズの堆積物中に鉛直に突き立ち、半分体を埋めた姿勢で生息している。シロウリガイは、自らの生存のために、体内に共生する独立栄養細菌に増殖に必要な酸素と硫化水素を供給しなくてはならない。前者は底層水中に含まれるものを水管経由で取り入れているが、後者は、堆積物の下層にあり、その摂取のために、体前端部を地中深く沈める必要がある。
 | 20-5 シロウリガイ(Calyptogenasoyoae)の生息状況。相模湾初島沖、水深1200メートル。 |
シロウリガイを解剖すると鮮血色の体液が流れ出ることからも明らかなように、この貝は呼吸色素ヘモグロビンを大量に持っている(太田ら1987、Suzukietal., 1989)。シロウリガイは、この血色素を用いて共生細菌に酸素を供給する。ところで共生細菌が必要とするもう一つの物質、つまり硫化水素は、ヘモグロビンと酸素よりも強力に結合するので、もしシロウリガイが単純に体内に硫化水素を取り入れようとすると、たちまち呼吸困難に陥ってしまう。シロウリガイでは、この問題を解決するために、体内に取り入れた硫化水素を運搬する専用のタンパク質を持ち、異なる生体高分子が分業して2種の物質を輸送すると同時に、細胞呼吸レベルでも硫化水素の解毒機構を備えている。
大部分のシロウリガイ類は巨大である。浅海の物理的撹乱の大きな場では、素早い掘削能力をもつか、厚い殻で比重を高め、堆積物上に安定して着座したり水流による洗い出しに耐えることも適応の一形態であろう。しかし、深海の軟泥底に生息する二枚貝は一般に薄質の殻をもち、殻長10ミリ前後と小型で、表面積/体積比をできるだけ大きくして、堆積物中で自重による埋積を小さくし、また限られた栄養で生活しようとするのと対照的である。シロウリガイ類は、場所にめぐまれれば生活に必要なエネルギー総量は十分であり、CCD以下の深海であったとしても、素材としてのカルシウムおよび炭酸の供給にも不足はない。堆積物に埋没しあまり水平運動をしない二枚貝の通性として、膨れた殻も不都合ではない。巨大な殻の空間は異常に分厚い鰓を収納できる。殻の厚み、すなわち強度の増大は耐捕食効果も高い。
多かれ少なかれ湧出孔生物は日和見的な種であり、密集型の幼生定着も考えられるが、分布の空間的局限性は、地下からの栄養供給路の空間的限定性、すなわちある閾値以上の濃度で希釈と酸化をうけない燃料物質の供給があり、酸化剤としての海水溶存酸素を同時に入手できる湧水−海水インターフェースにのみ限定されることにある。当然そこでは場所をめぐる激しい競争が行われる。密集による第二の効果は、繁殖の容易さであろう。一般的な深海底では雌雄邂逅のチャンスの確保は重大な関心事であるが、湧水生物群集においてはこの問題を免れている。近傍の個体と交尾する繁殖様式をもつ蔓脚類も熱水噴出孔において主要な構成メンバーとなりうるのである。
注目すべきは日本周辺におけるシロウリガイ類の種多様性である。これは調査密度、分類学レベルの高さを示すのみならず、日本周辺のテクトニックセッティングと地史の複雑さによるものであろう。むしろシロウリガイ類の生息が、冷湧水や熱水噴出孔などのテクトニックセッティングの指標となっている。また、深度別にすみ分けていることも気付かれる。たとえば、相模湾から銚子沖の数100〜700メートルにはアケビガイ (Akebiconcha kawamurai)、相模湾の初島沖・沖ノ山堆・三崎堆の750〜1300メートルにはシロウリガイ (C.soyoae)、駿河湾から遠州灘の1000〜2000メートル付近にはスルガシロウリガイ (C.fausta)、南海トラフ(遠州灘)の2000〜4200メートルにはテンリュウシロウリガイ (C.laubieri)、カイコウシロウリガイ(C.kaikoi)、ノチールシロウリガイ (C.nautilei)、そして日本海溝・千島海溝(〜オレゴン沖?)の陸側斜面にはナギナタシロウリガイ (C.phaseoliformis)が生息している。また拡大を開始しつつある沖縄舟状海盆には伊是名いぜな海穴、伊平屋いへや海嶺あるいは南奄西みなみえんせい海丘などの現在活動中の熱水噴出孔域にはエンセイシロウリガイ (C.solidissima)と未記載種 (Calyptogenasp.) が生息している。
遠州灘の2000〜4200メートルでは、同一属の数種が同一パッチに混在するという特異な事実に注目される。ほぼ同じ生活型の近縁種が同所的に優劣なく混在することは、生態学の常識になじまない。共生微生物や代謝経路を異にするなど思わぬすみ分けが起こっているのか、個々の環境の不安定さが大きく、生態学的な競争と淘汰による安定期にいたっていないと見るべきか、いずれにせよ将来の懸案となっている。なお、日本周辺のシロウリガイ類の遺伝子解析、アイソザイム分析による進化系統学は東京大学海洋研究所海洋生物生態部門で継続されている。また、狭義のシロウリガイも分子的には2タイプからなることが明らかにされ、分類学的再検討がまたれている。
シロウリガイ類のエネルギー基盤で述べたように、高密度で生息するかれらの生活を支える基幹物質は地下から供給されている。熱水域であれば熱水循環セルの開口部、沈み込み帯の遺留水湧出であれば断層面、高有機物含量の地下水の湧出であれば地下水脈の露出、石油・天然ガスのシープという裏付けをもたねばならない。その場合、硫化水素が海底下から直接供給される場合には(温度が許すかぎり)直接にその噴出に浴し、メタン等の低分子炭化水素の浸出であれば、低分子炭化水素−硫化水素カプリング層を形成するに適切な空隙率と透水率をもった比較的薄い堆積物層が必要条件となる。特に熱水噴出孔環境の場合、猛毒の硫化水素雰囲気への生理適応が要求され、通常の生物は厳しく排除される。熱水噴出孔および冷湧水生態系のもう一つの大きな特徴は、それぞれの環境が時空間的に不安定であり、高擾乱環境に数えられることである。個々の熱水噴出孔の寿命は短く、自らの噴出・沈着物質でその噴出孔を閉じてしまう。210Pbなどによる実測、長期観測による結果では個々の噴出孔の寿命は数十年の桁であり、多くの生物の寿命とほぼ同じ桁である。この3条件に合致すべく適応進化した化学合成生物群集は、必然的に厳しい隔離条件におかれる。さらに加えるならば、化石種とその産状を検討したり(横浜防衛施設局(編)1993、蟹江ら1992)、海溝軸・トラフ軸のジャンプや、伊豆半島(伊豆火山島)が本州弧に衝突し、駿河湾と相模湾を分断したような地史的イベントも加えるべきであろう。現在の日本周辺の西太平洋域に多数の冷湧水生態系や熱水生態系が存在するとはいえ、それらの個体群は、短いものでも数10キロメートル、多くは数百キロメートル、千数百キロメートルのオーダで互いに分断されており、これらの個体群がどのような適応分散と種分化の経路をたどって現在のような広い範囲に分布するようになったのかは、生物地理学の側面から興味深い。分子生物学的な手法から、各個体群の系統関係を解析した小島らの研究によると (Kojima et al., 1995)、南海トラフでは比較的遠縁の種が同所的に出現したり、同種あるいは姉妹種が沖縄トラフと相模湾に出現するなど、地理的に近い個体群が系統的にも近縁であるとはいえず、その適応放散の歴史は単純ではないようである(挿図6)。分子系統学に、地史的イベントと対応した絶対時間スケールをもたせる試みが進行中である。
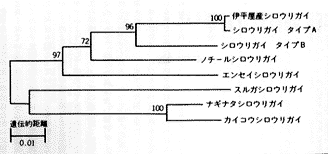 | 20-6 わが国周辺に分布するシロウリガイ類の系統関係 (Kojima et al., 1995) 。ミトコンドリアDNA COI, COIII 領域の同義的塩基置換に基づく、近隣接合法による分子系統樹。分岐のブートストラップの値が70%以上のものは、値を数字で示した。 |
以上、研究が進んでいる西太平洋域のシロウリガイ群集とその環境を中心として、化学合成生態系が動く地球とダイナミックにリンクしている例を紹介した。ただ、日本周辺では大型で目立つ生物であることと、硫化水素酸化型共生細菌に大部分の栄養を依存するシロウリガイが多く語られるが、現在の知識ではかれら、ならびにかれらの適応戦略は化学合成生物群集の一側面を語るにすぎない。世界の各地に発見されている熱水生態系や冷湧水生態系の化学合成生物群集は、シロウリガイ類、ハオリムシ類、シンカイヒバリガイ類、シンカイコシオリエビ類およびイトエラゴカイ類を主要構成要素とするものが多い。しかし、マリアナ背弧海盆、中央マヌス海盆、フィジー海盆、ラウ海盆では、エネルギー的に大きな鍵を握る両雄であるシロウリガイ類(硫化水素酸化型共生細菌依存型)とシンカイヒバリガイ(通常の懸濁食とメタン酸化型細菌との共生の両面作戦型)のニッチェを、なんと大型巻貝のアルビンガイ(Alviniconcha hessleri) やヨモツヘグイニナ(Ifremeria nautilei)が担っている。
また、熱水噴出孔環境は中央海嶺型拡大軸や背弧海盆リフト系中軸谷に限らず、島弧性火山フロントや鹿児島湾内のように水深百メートル以浅に吹き出す温泉でもハオリムシの密集群集がみつかったり、中生代や古生代の熱水噴出孔生物群集化石が発見されるなど、研究には数10キロメートルから中央海嶺系までの空間距離、0〜6300メートルの水深範囲、種々の地学的背景、個々の生物の寿命から地球の進化史までのスペクトルをもたなければならなくなった。あるいは、地球惑星の形成と生命の起源過程における前生命物質の合成に熱水噴出孔のような高熱還元環境が寄与したかもしれない。
化学合成生物群集の生物量は、通常の深海底のそれを四桁ほど上回るものであった。そして、シロウリガイ・シンカイヒバリガイ・アルビンガイ・ヨモツヘグイニナなどの主要構成生物である大型軟体動物の殻に取り込まれる炭酸カルシウム量は厖大である。湧水の停止でそれらの死殻が大量に堆積した墓場もよく見かける。また、冷湧水の場合、地中から浸出した低分子炭化水素は結局は二酸化炭素もしくは炭酸カルシウムとして生物体や殻、さらには地中に固結した炭酸カルシウムの導水管構造として残存する。地球上における二酸化炭素のトラップ形態として、その定量評価も重要な研究課題である。
化学合成生物群集の構成はたがいに驚くほど良く似ている。共生する独立栄養細菌についての分子生物学的な研究は、共生関係の進化が何度も起こっていることを示唆しているが、このような複数の種が関与する生物現象がどのようにして進化してきたのか、その進化の機構はダーウィン流の進化論で説明可能なのかなど、様々な興味深い研究テーマを我々に与えてくれる。かつて「種の起源」が執筆されるに当って重要な役割を果たしたガラパゴス諸島は、再びその沖合に熱水生態系という新たな現代生物学発展のきっかけをもたらしてくれた。
(太田 秀・白山義久)