




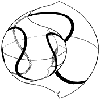

ニッポニテス~ラビリス Nipponites mirabilis Yabe
軟体動物門頭足綱アンモナイト亜綱
北海道留萌郡小平町達布
矢部久克
完模式標本UMUT MM 7560
総合研究資料館、地史古生物部門
ニッポニテスの螺環の巻方は、一見非常に複雑に見えるが決して不規則ではない。成長の初期には、殻は巻の解けた平面螺旋状かわずかにヘリコイド状(巻の軸方向に変位がある螺旋巻)を示している。成長の中期になると、初期殻の周りを蛇行しながら螺環が形成されていく(挿図1)。ちょうど野球のボールの縫い目のような図形を思い浮かべれば良い。ただし、ほぼ3回の蛇行を繰り返して螺環は巻き軸の周りを一周する(ボールの縫い目の場合は2回で一周する)ことから、矢部はこの巻方について「立方体の各面に沿うように螺環が成長していく」と説明している。後期になると蛇行螺環がやや乱れてくるか種によってはレトロバーサルフックと呼ばれる鈎状の住房を形成して成長を止める。
生活様式についても様々な憶測がなされてきた。古くは、ちょうど蛇がとぐろを巻いたような蛇行螺環が、いくぶん、現生のムカデガイ類(腹足綱)の殻形態に似ているので固着性の生活様式が推定されていた。また、海綿などの中に埋もれて生活していたのではないかという考えがあるが、化石の産状からはこの考えを支持するような証拠は認められない。最近では、ニッポニテス類もまた、他の異常巻アンモナイトや正常巻アンモナイトと同様に、海水中に浮遊するためにほぼ十分な体積の気房を発達させていたことが指摘され、浮遊生活者であったとする見解が優勢であるが決定的な証拠はまだ示されていない。
ニッポニテス属は、今日までに3種一変種が記載されており日本の他、サハリン、カムチャッカ、およびオレゴン(アメリカ)にまで分布し、北太平洋地域の白亜紀チューロニアン〜コニアシアンの時代を特徴づける。最近の研究によれば、北海道およびサハリン産のニッポニテスは、巻の様式や殻表面の装飾について互いに明確に区別できる3つの形態型が認められることが分かってきた(この3型のうち2つは、従来種のミラビリスおよびバッカスに対応するが第三の形態型については種名は与えられていない)。これらは時代的にも区別できるため、ニッポニテスが時間とともにわずかずつではあるが段階的に形態を変えてきたと思われる。矢部が記載したニッポニテス・ミラビリスは、本属の中でも最も古く(チューロニアン中期)原始的な形態型に含められる。この形態型の持つ成長初期の殻形態と、成長を通じての表面装飾の変化パターンは、矢部によって同時に記載されたユーボストリコセラス・ジャポニカムに酷似している(挿図2)。そのため、ユーボストリコセラス・ジャポニカムかそれに極めて類縁の種からニッポニテス属が派生したことはほぼ明らかであるが、成長の中期を通じて右巻あるいは左巻のヘリコイドを単調に持続するユーボストリコセラスと、螺環が激しく蛇行を繰り返すニッポニテスとの間の中間的な形態は発見されていない。
 |  |
| 18-1 成長の中期におけるニッポニテスの巻き方 | 19-2 Nipponites mirabilis YabeとEubostrychoceras japonicum(Yabe)の形態の比較。初期殻の形態と成長を通じての表面装飾の変化パターンは酷似しているにもかかわらず、中期以降の殻の巻き方はまったく異なっている。 |
(岡本 隆)
 |  |
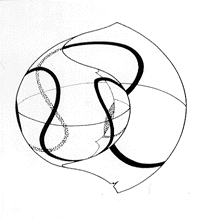
| 19-3 成長方向調節モデル(Okamoto, 1988)。蛇行螺環は生息姿勢をある一定の範囲内に保つための動的平衡機構の結果と考えられる。図は生息姿勢の範囲を0〜40』に設定したときの成長過程を模式的に描いている。 | 19-4 コンピューターによって再現された Nipponitesmirabilis(前)と Eubostrychocerasjaponicum(後)の理論的殻形態。表面装飾に関しては実際の標本を参考に人為的に描いている。 |