





Carcharias taurus
脊椎動物門軟骨魚綱板鰓類
東シナ海
谷内透
総合研究資料館、水産動物部門
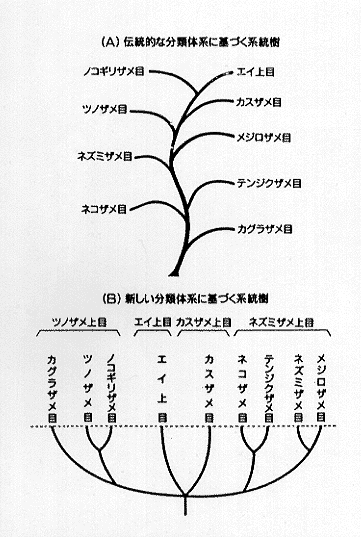 27-1 板鰓類の系統樹(谷内、1990)
27-1 板鰓類の系統樹(谷内、1990)
現今の系統分類の考え方の正否はさておき、現生のサメは分類上脊椎動物亜門の軟骨魚綱の板鰓亜綱 (Elasmobranchii) に属する。現生種は全世界に約350種、日本近海には約110種が分布する。硬骨魚に比べて種数が極めて少ないにもかかわらず、水平的には熱帯から寒帯まで、また垂直的には表層から数千メートルの深海にまで広い範囲に生息することが特徴である。大きさもわずか20センチで成熟する小型の個体から15メートルを越す巨体のものまで実に多様性に富んでいる。このように多種多様なサメの分類は必ずしも容易ではない。その理由の1つとして、サメには分類形質が乏しいことがあげられる。体形、鰓孔数、臀鰭の有無、各鰭の相対的な位置や形、頭部の形、眼や口の形や位置あるいはそれらの付属形質、体色などがキーとして用いられるが、明確に区分が困難な形質も多く、種の同定に際ししばしば問題を提起する。また、頭骨は1つの脳函からなっていることから、硬骨魚のように内蔵骨の構造に基づく分類が極めて困難であるという弱点も有する。しかも内部骨格は軟骨性であることから化石として出現することは極めて稀であるため、化石に基づく統類縁関係を模索することは不可能に近い。このため、現存種の骨格系や筋肉系の構造から、系統類縁関係を構築することが行われているが、古生物学的な裏付けがないことから、化石種と結びついた体系を打ち立てにくい。したがって、一般的に現生種に基づく分類体系と化石に基づく系統分類との間に乖離が起きやすく、しばしば論争の種となる。
サメの歯は次々に生え変わる多生歯性であり、機能歯の舌側には常に数列の歯胚が存在する。歯の交換様式は車軸交換と呼ばれる。そのメカニズムは後藤 (1985) によれば、次の通りである。まず歯の硬組織が完了する頃になると、歯胚は外側に回転して露出し機能歯(作用歯)となる。機能歯はさらに外側に移動して唇の外縁部にある陥没部に達すると、歯根部が上皮中に押し上げられ、歯は脱落していく。理解を容易にするため、サメ下顎縦断面の模式図を示した(挿図3)。 この模式図では機能歯は唇上に2本が露出し、また最上部の置換歯は 口腔粘膜上皮から萌出するが、その下にある三本の置換歯は口腔粘膜上皮に保護されている。唇上にある歯が脱落すると、下方(舌側)の置換歯がエスカレーター式に回転しながら唇方向に移動し、機能歯となる。一般に歯は口の奥の方に傾く。ニシレモンザメ (Negaprion brevirostris) を用いた飼育実験では、上顎の歯は平均7・8日、下顎の歯は平均8・2日で生え変わる (Moss, 1967)。しかし、成魚になると磨耗した歯が数多くみられることから、換歯率は若いときほど早くないものと推定される。また、ニシレモンザメを含むメジロザメ目やネズミザメ目のサメでは、それぞれの歯は単一に脱落することが多いが、隣接する歯の歯根部が接する歯列を持つツノザメ目のサメでは、歯は一列同時に脱落する。後藤 (1985) は、サメ類の歯は未分化な硬組織からなるという原始性を保持する一方、車軸交換という特殊な交換様式を発達させていることから、原始性と進化性を合わせ持つ特異な生物としている。確かに、このような交換方式の歯を持つことは、捕食者としての地位を確立するのに有効である。
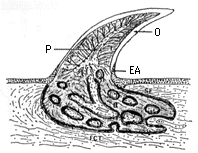 | 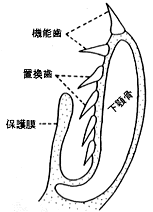 |
| 27-2 歯の組織構造(後藤{、1976) O:エナメル質(エナメロイド)、 I:象牙質、B:歯根部の骨様組織、 P:歯髄、 EA:付着上皮、 SF:シャーピー線維、 FCT:線維性結合組織層。 | 27-3 サメの下顎縦断面(Cappetta, 1987) |
このような顎全体の歯の形や数とともに、1本の歯の形態が特に古生物の分野で重要である。というのも、軟骨魚においては化石として現出するものはほぼカルシウムが沈着した歯に限定されるので、1本の歯のかなり微細な形の違いに基づき新種の記載がしばしば行われるからである。したがって、歯の形態を記述するには各部位に統一的な名称が必要である。しかし、残念ながら各部位の名称は必ずしも統一されているとはいえず、研究者により独自の名称が使われることがある。ここでは、後藤 (1978) が提示した名称に従うことにしよう(挿図5)。まず、歯は歯冠 (crown) と歯根 (root) に分かれ、両者の境目は歯頚 (neck) といわれる。歯冠にはふつう中央部が突出した咬頭(現生種の分類ではよく尖頭という言葉が使われる)があり、通常は1個であるが、時には複数の咬頭を持つ種類もある。この多咬頭のうち、最大のものを主咬頭、小さいものを副咬頭と呼んで区別する。副咬頭の有無やあるいはその数が重要な分類上の手がかりとなることがある。たとえば、ネズミザメ科では副咬頭を持つ属はネズミザメ属だけであり、他の形質が類似していても、歯をみれば容易に識別できる。また、シロワニとオオワニザメ (Odontaspis ferox) の主要な区別点は、前者が通常はそれぞれの歯の両側に一副咬頭を持つのに対し、後者は通常は二副咬頭を持つことである。咬頭の切縁部がのこぎり状の形状を呈する形態(鋸歯)も重要な分類形質となる。特にメジロザメ科では鋸歯縁を持つかどうかが属の検索に用いられている。すなわち、下顎歯が鋸歯縁を持てば、メジロザメ属に行き着き、さらに上顎歯が鋸歯状であるかどうかも種の検索において重要な手がかりとなる。さらに現生種の分類では歯の基底部に深い欠刻がある(挿図5では切痕という名が付けられている)ことも種を分ける基準に用いられる。挿図5にみられるようにイタチザメは深い切痕を持つことで特徴づけられる。もちろん歯の全体の形状、すなわち、牙状であるか、三角形を呈するか、咬頭が傾き隣接の歯と接触するか、また臼歯状であるかはサメの目または科を特徴づける形質として重要である。
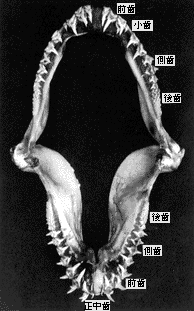 | 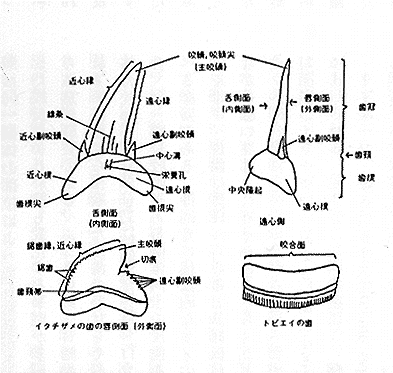 |
| 27-4 シロワニの上{にある歯の名称。前歯、小歯、側歯、後歯、正中歯[原図](Taniuchi, 1970) | 27-5 歯の各部位の名称(後藤、1978) |
サメでは皮歯の形状や構造が時には重要な役割を演じていることがあり、外形や歯とともに図示されることが多い。ただし、皮歯の形は同一個体の部位によっても変異があり、また成長により形態も変化する事例が多いから注意が肝要である。ふつうは第一背鰭起部直下の側面から採取した皮歯の形状をその個体の代表とする。また種間の比較を行う場合、同一成長段階にある個体同士の皮歯を比べる必要がある。挿図6に代表的なサメの皮歯を示す。皮歯は側面からみると基底板につながる細い茎部上に冠部がある。ちょうど燭台のような形を思い浮かべればよい。上方からみると葉状を呈するものが最も一般的で、時には槍形あるいは角形を呈する。多くのものは葉状の冠部に隆起線が3本または5本縦走する。ビロードザメには縦の隆起線に加えて数本の横走する隆起線が走る。カラスザメ属では剛毛状の皮歯を持つものがある。また、ユメザメでは円形の冠部の中央が凹み、皮膚をなでると傷つくことがある。
皮歯の役割については4つの機能があげられている (Reif, 1985)。第一の機能として、 皮歯は寄生虫を含む外的からの攻撃に対する防御として役立つ。この例としてはツノザメの背鰭棘やキクザメの瘤状皮歯が挙げられる。第2として、底生生活をするサメの皮膚を防護する機能である。ネコザメやカスザメの腹側の皮歯にはよく鋭いひっかき傷や擦れがみつかるが、これは岩や石に接触する機会の多いこれらの底生性のサメにとっては、皮歯が表皮を防護する機能を持つことが容易に推察されるだろう。第3に、高速で遊泳する外洋性のサメの皮歯は密に重なり合うと同時に冠部には高い隆起と深い溝があるため、抗力や渦流を減ずる役割があることが指摘されている。第4に、皮歯は発光器と密接な関係にあり、生物発光に貢献していることがあげられる。カラスザメ、カスミザメ、ダルマザメの体側や腹部には多数の発光器が皮歯の配列と密接な関係を持つ。
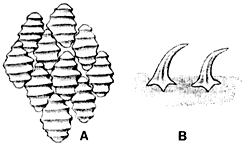 | 27-6 サメの皮歯(谷内、1976)。A)ヨゴレ Carcharhinuslongimanus、B)フジクジラ Etmopteruslucifer。 |
 | 27-7 代表的な歯の形状。A-Dは BigelowandSchroeder(1948)より、Eは谷内(1976)、FはCappetta(1987)より引用。A)噛み付き型(トラザメ類 Scyliorhinusretifer)、 B)引き裂き型(アオザメ Isurusoxyrinchus)、C)純切断亜型(アブラツノザメ Squalusacanthias)、D)切断氓沍^(カグラザメ Hexanchusgriseus)、 E)押し潰し型(ホシザメ Mustelusmanazo)、F)すり潰し型(ネコザメ属 Heterodontus) |
まず第一に、噛み付き型 (clutching type) があげられる。基本的には同形歯型で、時には部位によりやや形が異なる。歯は小さく、多くの場合複数の副咬頭を持ち、餌の捕捉をより容易にできるような構造となっている。歯は数列が同時に機能し口角部と内側に向かって傾いている。この型の歯を持つサメは、トラザメ科、ドチザメ科、テンジクザメ科のような沿岸性や深海性の小型魚に多い。次に引き裂き型 (tearing type) がある。この型のサメの歯は、ほっそりした主咬頭に時には数個の副咬頭があるが、全くこれを欠く種類も多い。一般にほっそりした歯の両縁辺は鋭く、獲物を引き裂いたり引きちぎったりするのに好適である。機能歯は数列並び、それぞれの歯はやや口角部に傾くと同時に鋭く口の奥にカーブする。後歯は小さく、一見臼歯状を呈する。ネズミザメ科ヤミツクリザメ科さらにシロワニ科のサメはこの型の歯を持つ。第3に切断型 (cutting type) がある。この型はさらに2つのサブタイプに分けられる。純切断亜型 (cutting sensu strico subtype) は、口角部に向かいわずかに形が小さくなるが、ほぼ同形歯型を示す。歯の幅が広く、主咬頭は口角部の方向に鋭く傾く。隣接する歯は互いに重なり合うか密に接触する。機能歯は一列が原則で、歯は一列同時に脱落する。典型的な例はツノザメ類で、その変形がイタチザメにみられる。もう1つのサブタイプは切断・押し潰し亜型 (cutting-clutching subtype) で、歯はむしろ異形歯性を示し、部位により歯の形状は著しく異なる。典型的な例がカグラザメで、上顎は前歯では主咬頭のみ、側歯になると一副咬頭が生じ、その数を増していくが、後歯ではむしろ臼歯状に近い形状となる。下顎歯では、まず正中歯は左右相称に近く両側に複数の副咬頭があるが、前歯から側歯にかけてはやや大きな主咬頭から順次小さくなる多数の副咬頭が口角部側にみられる。後歯は上顎同様臼歯状となる。純切断亜型に比べてこの型の歯は獲物を切断して噛み込むのに有効な様式の歯を持っているといえよう。このほか、メジロザメ科のサメでは前方では噛み付き型で口角部に近くなると臼歯状の歯が発達するものが多く、採餌にきわめて有利な歯を持つことが分かる。第4に、押し潰し型 (clutching type) の歯がある。歯が尖らず平らであることが特徴で、獲物を押し潰すのに適応している。モザイク状に並んだ機能歯が4〜5列露出する。貝や頭足類を押し潰して採餌するのに最適な歯である。エイ類の多くはこの型の歯を持つが、サメではホシザメ類がその典型である。この変形として、歯が癒合して強固な1枚あるいは複数の歯板を形成するすり潰し型 (grinding type) がある。 その典型はギンザメであり、また板鰓類ではトビエイ類がこの型の歯を持つ。最後に押し潰し・すり潰し型 (clutching-grinding type) があげられる。ネコザメにみられるように、前歯は捕捉型、側歯や後歯はいくつかの歯が癒合して歯板型となる。前方で獲物を捉え、後方でかみ砕いたり咀嚼したりするのに有利な歯型である。ちなみにネコザメはサザエ割りの異名がある。
歯とともに顎の筋肉や骨格型と関連させ、捕食機構からみた進化の図式も提出されている (Moss, 1977)。この説を谷内 (1988) に従い説明しよう。古生代型のサメは長い吻の 先端に口がつき、さらに顎の懸垂法も頭蓋骨と2ヶ所でつながる両接型であり、歯も碇型であったことから、まず初めに獲物を捕捉するのに便利な把掴型の捕食様式が発達したのだろう。やがて顎の懸垂法も舌顎軟骨を介して頭蓋骨とつながる舌接型となることにより咬む力は増大し、口もすぼんだため吸引-把掴型に進化した。吸引・把掴型からは3つの型が派生したと考えられる。1つはツノザメ類にみられるように切断型となり、他はトビエイ類にみられるような吸引・咀嚼型、もう1つはネズミザメ類やメジロザメ類にみられる丸呑み型になった。丸呑み型では概して大きくて鋭利な歯をもち、しかも歯数が少ないという特徴がある。胃内容物を調べると、この型の歯を持つサメからは丸のままか大きすぎれば2つか3つに切断された餌生物が見つかる。丸呑み型からはさらに2つの型が分化し、1つはドチザメにみられるような咀嚼型に、他はテンジクザメ類にみられる吸引−咀嚼型になった。ジンベイザメやイトマキエイにみられる特殊な捕食様式である濾過型は、トビエイ型とテンジクザメ型の吸引−咀嚼様式および丸呑み型から発展したものと推測している。Moss (1977) は板鰓類が海洋における多用な餌環境に適応した結果、このような様々な捕食様式を発展させたものと考えている。
まず歯の配列様式からみたサメの進化を、後藤・橋本 (1976) の説に従って説明しよう。デボン紀に出現したクラドセラケ (Cladoselache) は長い顎に幅広い間隔の歯族が並ぶことから、最も原始的な配列様式と考え、現生種のラブカにみられるのでラブカ型と名付けた。現代型のサメの中ではネズミザメ類が比較的長い顎を持ち、並列型の配列様式を持つことから、次の進化の段階と考え並列型と命名した。最も顎が短いメジロザメ類では、歯がモザイク状に並ぶことから、交互型と名付けた。顎の短縮が進化の指標とすれば、歯の配列様式もラブカ型→並列型→交互型と進化したものと考えている(挿図8)。
 | 27-8 歯の配列様式の進化(後藤、1985を若干改変して引用) |
次に歯の形態からみたサメの進化を考えてみよう(後藤1985)。クラドセラケやラブカにみられるような三咬頭性をまず歯の基本形に据える。この基本形から副咬頭数の増減の二分極化がみられ、五および七副咬頭に増加したヒボータス類と二副咬頭に減少したクテナカンタス類に分化した。一方、歯の形状そのものも食性の多様化により、臼状や板状の歯が生まれた。クラドーダス段階に出現した Protacrodus にその兆候が現れ、ヒボータス段階ではさらにこの傾向が顕著となり、現生のエイ類にみられる臼歯状の歯に進化したものと考えられる。現生のネズミザメ類では、ホオジロザメのような歯の巨大化がみられる一方、ウバザメやジンベイザメなどのプランクトンフィーダーにみられる歯の矮小化が起きている。肉食型の捕食様式も多様化して、その食性に応じて切縁歯、鋸歯などの肉食性に適した形態から有殻無脊椎動物を採餌するのに適した臼状や板状の形態までの間のスペクトルに様々な歯が位置している。挿図9に後藤の仮説を示した。
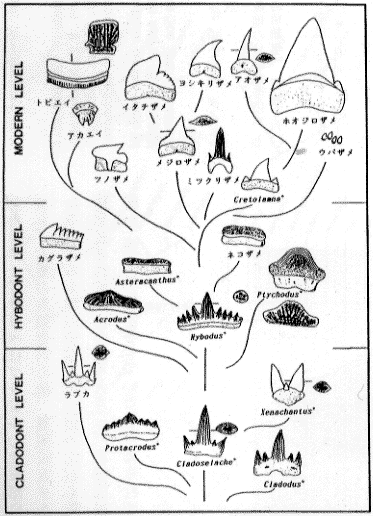 | 27-9 歯の形からみたサメの進化(後藤、1985を若干改変して引用) |
さらに、歯の組織構造からみた進化についての考察もあるが、詳細は後藤 (1985) を参照されたい。なお、かつては象牙質の組織構造の違いに基づき板鰓類を真正象牙質類 (orthodonts) と骨様象牙質類 (osteodonts) に2分することも試みられたこともあるが、現在では顧みられていない。
(谷内 透)