





Axelrodichthys araripensis Maisey
脊椎動物門硬骨魚綱肉鰭類シーラカンス科
ブラジル、セアラ州、下部白亜系、サンタナ層
約1億1000万年前
富永義昭
総合研究資料館、動物部門
このシーラカンスは、体長が1〜2メートルに達すること、第一背鰭条10〜11、第二背鰭条9〜10、尾鰭の上葉17〜18条、下葉は15〜16条、胸鰭条17〜18であること、頭蓋や角骨の表面が粗雑であること、主鰓蓋骨や喉板に多数の細かい放射状の線が走っていること、よく化骨した壁をもつ鰾が第二背鰭の後方まで延長し、くびれによって2ないし3室に分かれていることなどで特徴づけられる。シーラカンス類における本種の系統的位置については、いくつかの研究がある (Forey, 1988; 1991; Cloutier, 1991)。これは、現生種 Latimeria chalumnae の最近の系統分類学的研究によるところが大きい。挿図1は、 いずれも Axelrodichthys 属が Mawsonia 属と姉妹群を形成することを示している。これは両属が共通の祖先をもつことを意味する。Mawsonia 属には、アフリカの下部白亜系から4種、南アメ リカ・ブラジル・バヒア州の下部白亜系から一種 M. gigas, Axelrodichthys araripensis と同じブラジル・セアラ州の下部白亜系サンタナ層から産出する M. cf. gigas(M. gigas に近縁な種)の計6種が含まれている(CloutierandForey, 1991; Maisey, 1986; 1991)。このシーラカンスの仲間も浅海か淡水域に生息していたらしい(Maisey, 1991)。Mawsonia 属の分布は古生物地理学的に重要で、その種分化にはアフリカ大陸と南アメリカ大陸の分割が大いに関係していたと推測される (Maisey, 1986; 1991)。しかしながら、M. cf. gigas と同じ地層から Mawsonia 属に極めて近縁な Axelrodichthys araripensis が発見されたことは、Mawsonia 属の古生物地理と種分化の研究に再検討をせまることになるかもしれない (Maisey, 1986)。
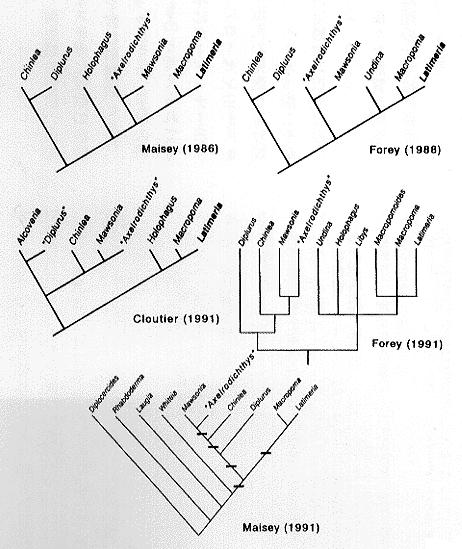 | 28-1 Axelrodichthys属のシーラカンス類における系統的位置。Maisey(1986)の系統樹は Cloutier(1991)より引用。 |
ところで、このシーラカンスで興味深いのは、鰾の保存状態である。本種の鰾は多くの他のシーラカンスの化石種と同様によく化骨しているが、他の種のように押しつぶされていない。つまり、膨らんだ状態で保存されている。本化石の鰾は堆積時の圧力に抗しており、このことは鰾が生時には脂肪に富んだ組織で満たされていたことを示唆している。さらに、このことは鰾内に堆積物がないことからも示唆される (Maisey, 1986)。
現生のシーラカンスは太くて長い鰾をもっているが、その中には水より軽い脂肪が充満している。壁は化骨していない。これは、海の表層と深海を移動する生活への適応と考えられている。鰾に気体が入っていると、水深により水圧が変わったときに体積を調節することが困難で、このような深浅移動をするような生活はほとんど不可能だからである(上野1992)。
ところが、モイセイ (1986; 1991) によれば、Axelrodichthys は浅海か淡水域に生息していたにもかかわらず、脂肪の充満した鰾をもっていたことになる。しかし、一般に鰾に脂肪を満たしている魚類は、現生のシーラカンスやハダカイワシ類のような深浅移動をする深海魚に限られており、本種のように浅海か淡水域に生息していたと考えられる魚類が脂肪を充満させた鰾を持っていたということは、これまでの鰾の構造と生息環境の関係(註1)からは理解し難い(註2)。
この点に関して、上野 (1992) は、膨らんだ状態で化石になっている鰾には気体が入っており、このシーラカンスが浅い海で生活していたのがその証拠であると考えている。この考えの方が、一般的な鰾の構造と生息環境の関係からみて合理的である。
しかし、本種の鰾が膨らんだ状態で保存されていることに関してのモイセイ (1986) の説明には説得力があるように思える。この問題については、これまでの鰾と生息環境の関係の見直しも含め、詳細に検討する必要がある。
シーラカンスは4足動物の祖先を考える上で重要な位置を占めてきた。それは両生類に始まる4足動物の祖先は総鰭類のシーラカンスやユーステノプテロンの仲間であると考えられてきたからである。
ごく最近に至るまで、両生類の直接の祖先は原始的両生類イクチオステガに近似の総鰭類ユーステノプテロンを含む扇鰭類とされ、この考えはジャルヴィック (Jarvik, 1942) の研究以来疑うべからざるものと考えられてきた。ところが1980年にガーディナー (Gardiner, 1980)、1981年にローゼン (Rosen, et al., 1981) らは4足動物の祖先の見直しを分岐分類学的方法により行い、ハイギョ類こそが両生類の祖先すなわち4足動物の祖先なのだということを主張した(上野1988)。
この論争においては、様々な形質が問題となっているが、大きな問題の1つは内鼻孔の解釈に関するものである。上野 (1988) によれば、1950年代まで内鼻孔をもつ内鼻孔類として総鰭類とハイギョ類がまとめられていた。しかし、現生のシーラカンスには内鼻孔がないことが判明し、化石のシーラカンス類にもなかったであろうと考えられるようになった。またユーステノプテロンにも内鼻孔はなかったのではないかとの主張もでてきた。一方、ハイギョ類の内鼻孔に関してジャルヴィック (1995) は後鼻孔が口の中へ移動したもので、4足動物の内鼻孔とは違うものであると主張したが、ガーディナー (1984) やローゼンら (1981) は後鼻孔が口中に移動したハイギョ類の内鼻孔こそが4足動物の内鼻孔と相同であると主張している。この考えに基づき、最近では内鼻孔類はハイギョ類と4足動物を含めたものとして使われている。
この論争は今日に至ってもまだ続いている。
(坂本一男)
註1 一般に、浅所を遊泳する魚類は気体を充満させたよく発達した鰾をもっている。