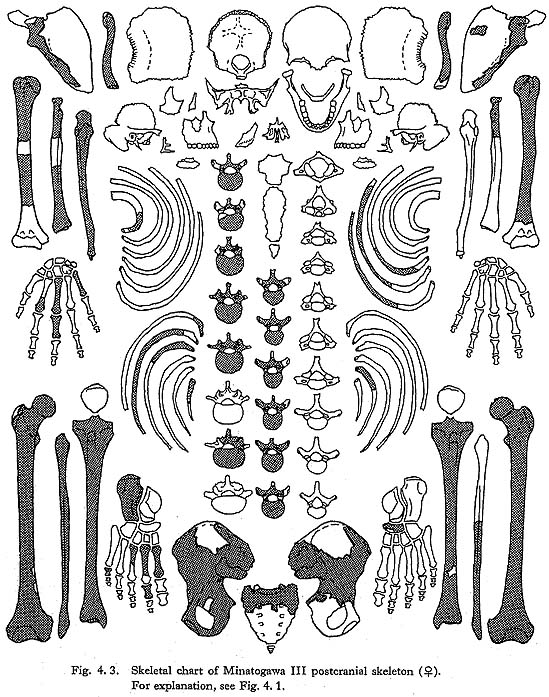
CHAPTER 4
Postcranial Skeleton of the Minatogawa Man
Hisao Baba* and Banri Endo**
*Department of Anatomy, Dokkyo University School of Medicine;
**Department of Anthropology, Faculty of Science,
The University of Tokyo
| ( 4 / 7 ) |
|



   |
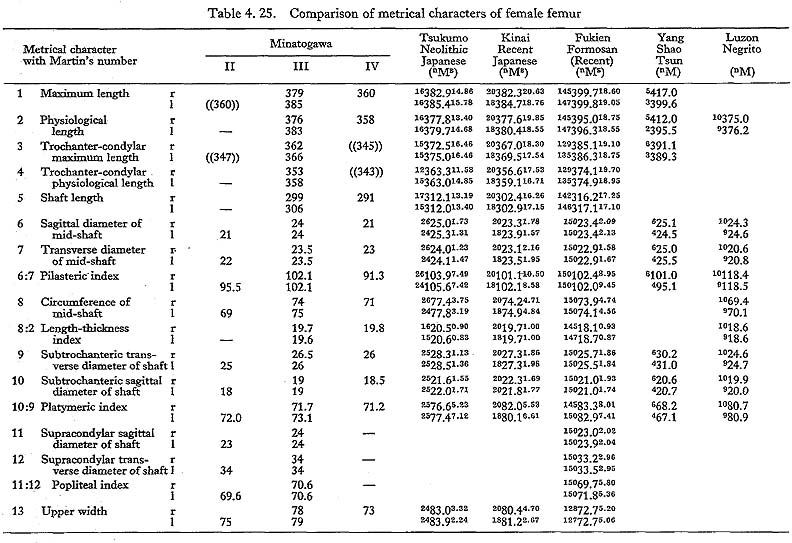   |
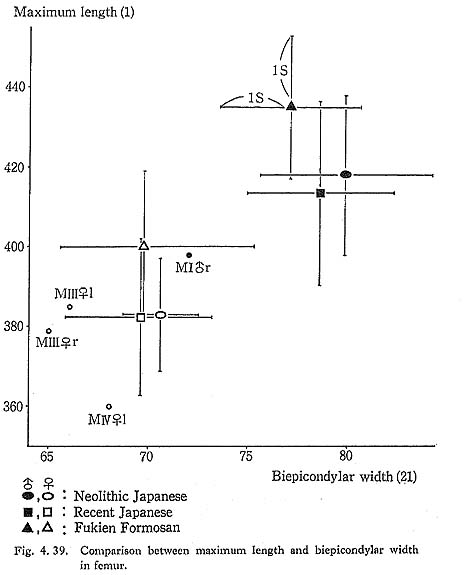
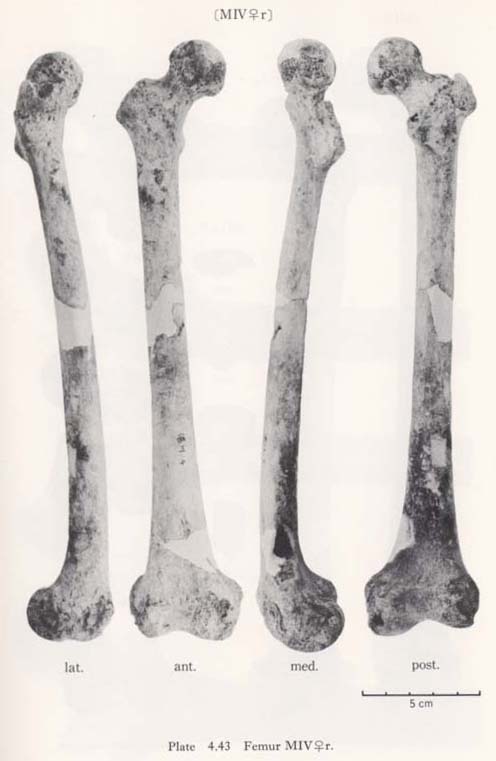
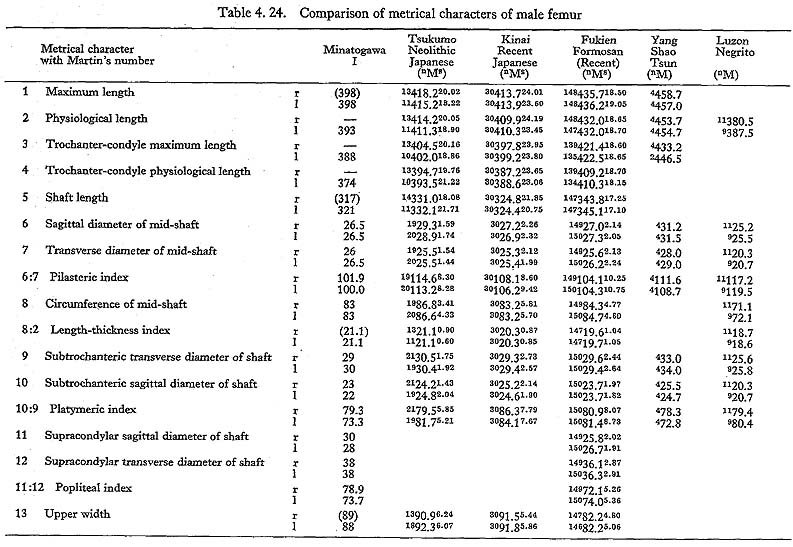
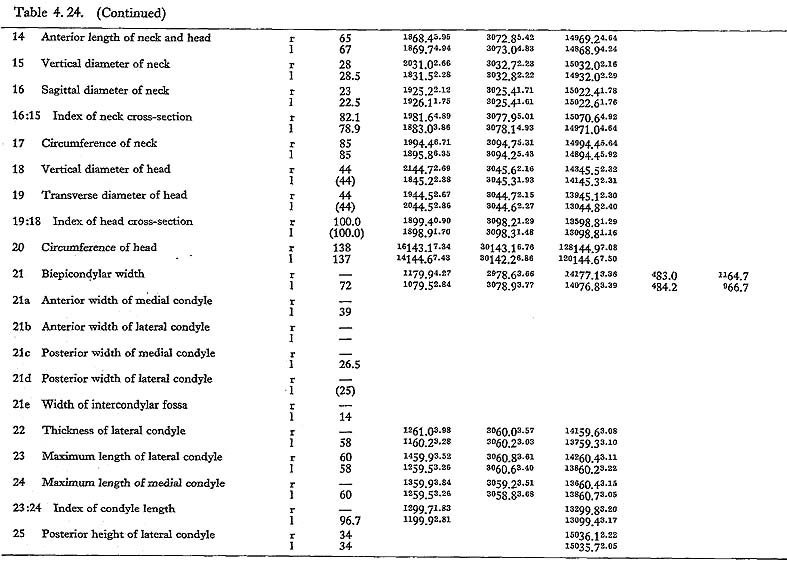
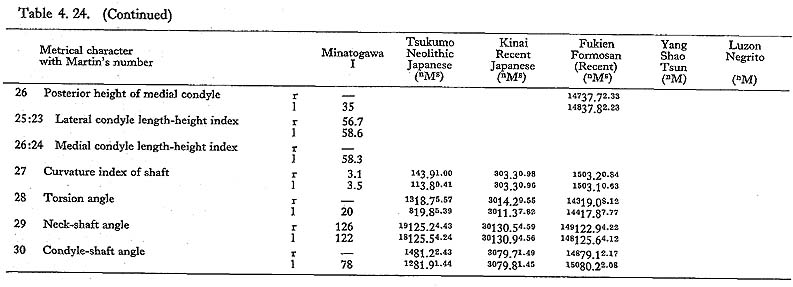
The maximum length of MIV r is also smaller than that for the Neolithic and Recent Japanese females. The estimated value in MIIl is also small. MI
r is also smaller than that for the Neolithic and Recent Japanese females. The estimated value in MIIl is also small. MI rl, MIIl and MIVr show, therefore, the same tendency in their length being considerably smaller than those for the Neolithic and Recent Japanese and Formosan Chinese or Neolithic Chinese but similar to Luzon Negrito. On the contrary, the maximum length of MIIIrl is comparable to the mean for Japanese populations.
rl, MIIl and MIVr show, therefore, the same tendency in their length being considerably smaller than those for the Neolithic and Recent Japanese and Formosan Chinese or Neolithic Chinese but similar to Luzon Negrito. On the contrary, the maximum length of MIIIrl is comparable to the mean for Japanese populations.
Proportion
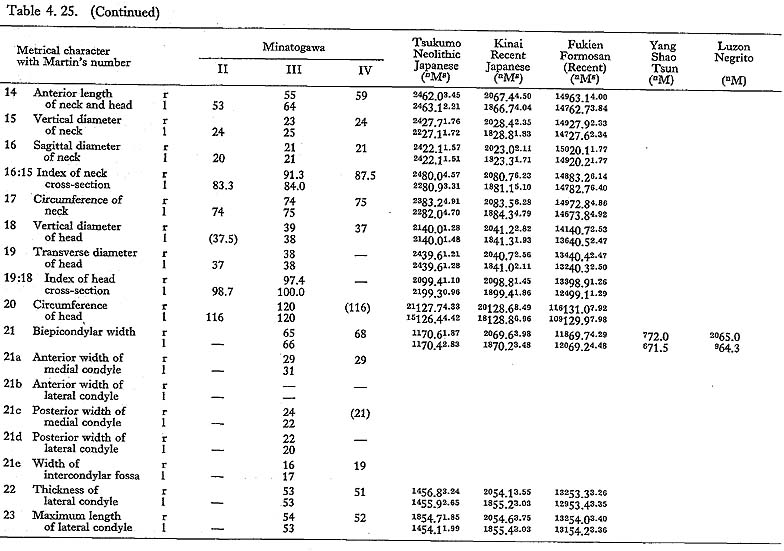
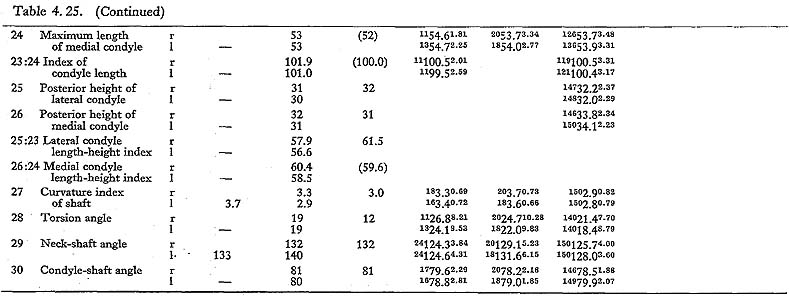
In MIr, the circumference at mid-shaft is 83 mm, and the length-thickness index is 21, being equal to the mean for Neolithic Japanese males. In MIIIl, the circum-ference is 74 mm and the index is 20, both of which are almost the same as the means for the Neolithic and Recent Japanese females. The circumference, however, is 69 mm in MIIl and 71 mm in MIVr, far smaller than those for the other populations. The indices (19 and 20, respectively) are rather close to those for the Japanese popu-lation but are larger than the mean for Negrito.
Neck-shaft angle
This angle varies from 122° to 140° in Minatogawa femora, which is almost equal to the ranges for the Neolithic and Recent Japanese (114°-146°).
Condyle-shaft angle
This angle is 78° in MIl, being slightly less than the means for the Japanese and Formosan males. It is 81° in MIIIr and MIVl, 80° in MIIIr, all of which are slightly larger than those for the Japanese and Formosan females.
Torsion angle of the shaft
This angle of MI is 20° in the left bone and roughly estimated as 13° in the right bone, both of which are within the range of all male populations compared considering the wide variation range. The case of MIIIrl is also similar. But in MIVr it is 12, which is smaller than that for the other populations compared.
Upper Epiphysis
Overall size
The upper epiphyseal length in MIl is almost equal to the mean for Neolithic Japanese males, and a little smaller than that of Recent Japanese males. In MIIIl, this length is close to that for the Neolithic Japanese females. These values in MIIl and MIVr are considerably smaller. In proportion to the maximum length, the upper epiphyseal length is great in MI and moderate in MII, MIII and MIV.
Head
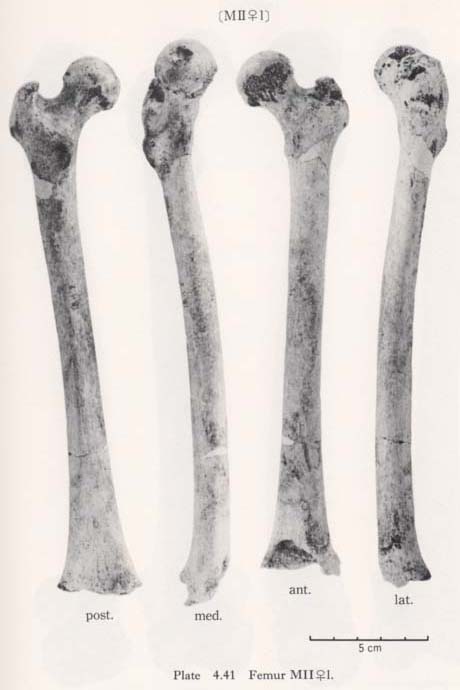
The circumferences of the heads in MIl and MIIIl is smaller than those for the Neolithic Japanese and Fukien Formosan. MIIl and MIVr show an equal value: 116 mm. The difference between this value and the mean for the females compared is large and about three times greater than the standard deviation for Neolithic Jap-anese females, i.e., they are extraordinarily small.
The Fovea capitis femoris is well defined by its sharp edge running from the head articular surface in MI and MII. In MIII and MIV, the edge is rather blunt. The fovea is generally deep but narrow, its maximum diameter being 14 mm in MIr, 12 mm in MIIl, 10 mm in MIIIl and 13 mm in IVl.
Neck
The most conspicuous impression on the upper epiphysis is a constriction of the neck in contrast to the usual development of the trochanter region. The vertical diameter of the neck of the Minatogawa specimens is far smaller than the means for other popu-lations compared.
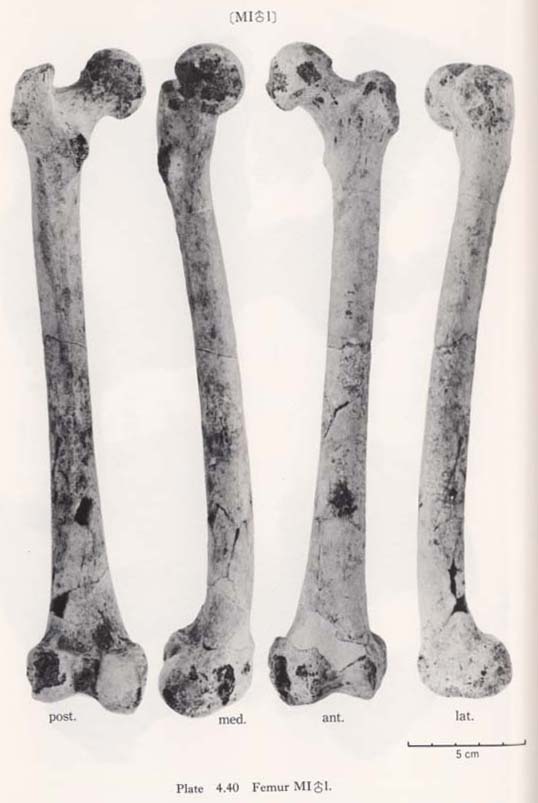
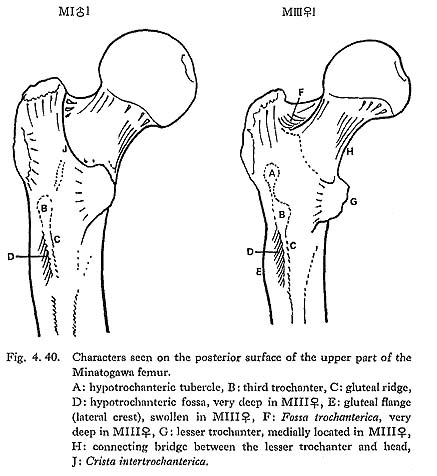
The so-called connecting bridge ( , 1930) from the lesser trochanter to the head is seen in MIIIrl and MIVr, but is not seen in MIrl and MIIl (Fig. 4. 40-H).
, 1930) from the lesser trochanter to the head is seen in MIIIrl and MIVr, but is not seen in MIrl and MIIl (Fig. 4. 40-H).
Generally, an imprint or roughness is sometimes seen on the antero-superior surface of the neck, which is called the "cervical fossa of Allen" (Charles, 1894a; Mayer, 1924) (Fig. 4. 41). In all these six femora this fossa is seen, but it is not as marked as in the Neolithic and Recent Japanese.
Trochanter region
The greater trochanter of Minatogawa femora is relatively well developed. Its height is 43 mm in MIl, 36 mm in MIIl and 33 mm in MIIIl. Its width is 35 mm, 32 mm and 33 mm in the same specimens, respectively. As compared with the Recent and Neolithic Japanese specimens, the width seems larger in proportion to the height in Minatogawa specimens.
The intertrochanteric line is seen in MIrl, MIIl and MIVr, forming an inter-mittent crest which separates the neck from the shaft. Sometimes the uppermost part of the crest increases the height to become a femoral superior tubercle (anterior tuberosity of the great trochanter, McCown and Keith, 1939). In MIrl, MIIl and MIVr this femoral tubercle is weak. Conversely, the line is scarcely visible but the tubercle is remarkable in MIIIrl.
The Fossa trochanterica is generally deep (Fig. 4. 40-F). In MIII the fossa extends onto the superio-posterior surface of the neck, which resembles the digital fossa de-scribed by McCown and Keith (1939). The so-called exostosis in trochanteric fossa (cf. Finnegan, 1978) is absent.
As for the Crista intertrochanterica, there are two types in Minatogawa femora. In MIl and MIIl it is well developed forming a high straight ridge from the greater trochanter down to the lesser trochanter (Fig. 4. 40-J). This crest in MIr is not ob-servable because of the breakage. In MIIIrl and MIVr, however, the crest runs downward from the greater trochanter and, after forming a quadrate tubercle, de-creases in height and becomes a low eminence continuing to the lesser trochanter. The former type is seldom seen in the Recent Japanese but is sometimes seen in Neolithic Japanese.
The lesser trochanter is generally well developed. In MIII it is remarkably devel-oped in contrast to the overall slenderness (Fig. 4. 40).
The lesser trochanter is posteriorly situated on the surface of the shaft in MIrl and MIIl, On the contrary, it is posteromedially in MIVr and medially in MIIIrl, having a close relationship to the connecting bridge described before.
There are typical third trochanter on the lateral border of the posterior surface at the lesser trochanter level in MIrl, MIIIrl and MIVr (Fig. 4. 40-B). In MIIl, the condition is not clear because of breakage.
In MIIIr the Tuberculum hypotrochantericum (Weidenreich, 1941) is well developed forming a round eminence (Fig. 4. 40-A). Its vertical and transverse diameters are 12 mm and 17 mm, respectively. This tubercle is absent in MIIIl and MIVr, but the upward extension of the gluteal ridge is seen. In MIrl there is also a weak extension of the gluteal ridge which runs not toward the greater trochanter but toward the intertrochanteric crest. In MIIl no tubercle nor extension of the gluteal ridge is seen.
Diaphysis
Form as a whole
The shaft of Minatogawa femora is generally straight. From an anterior view, the shaft is narrowest at the mid-point and becomes wider upward as well as downward. Seen from a side view, within the lower half of the shaft the sagittal diameter is almost equal, but from the mid-point up to the subtrochanteric region, the shaft gradually decreases in diameter.
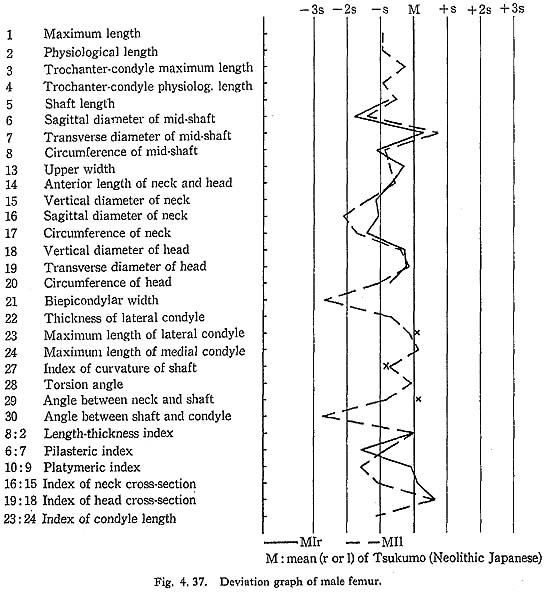
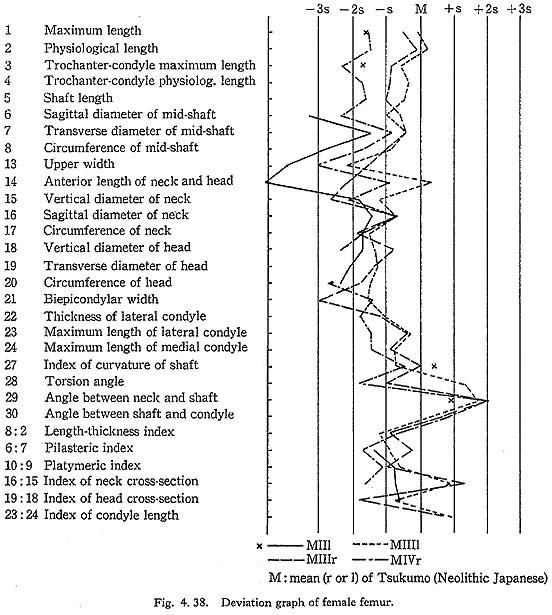
The curvature index of the shaft gives the range value of 2.5-3.5, which occupies the lower part of the range for Neolithic Japanese (2.1-6.9) and for Recent Japanese (1.5-5.2). The vertex of the curvature is situated distally in MIrl and MIIIrl and medially in MIIl and MIVr.
Subtrochanteric region
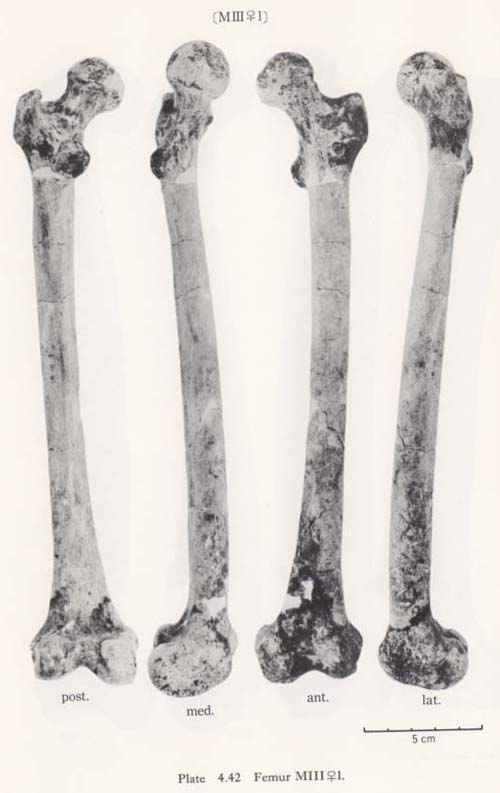
The real medial oblique ridge, as seen in Neanderthals as  Ea (McCown and Keith, 1939), in Amud I (Endo and Kimura, 1970) and in Fossil Sapiens as Obercassl (Bonnet, 1919), is not present in the Minatogawa femora. But a long ridge runs down along the antero-medial surface of the shaft from a position immediately under the neck and to mid-shaft forming a medial margin in all the present specimens (Fig. 4. 42-MC). This ridge is equivalent to the Crista medialis in Sinanthropus (Weidenreich, 1941).
Ea (McCown and Keith, 1939), in Amud I (Endo and Kimura, 1970) and in Fossil Sapiens as Obercassl (Bonnet, 1919), is not present in the Minatogawa femora. But a long ridge runs down along the antero-medial surface of the shaft from a position immediately under the neck and to mid-shaft forming a medial margin in all the present specimens (Fig. 4. 42-MC). This ridge is equivalent to the Crista medialis in Sinanthropus (Weidenreich, 1941).
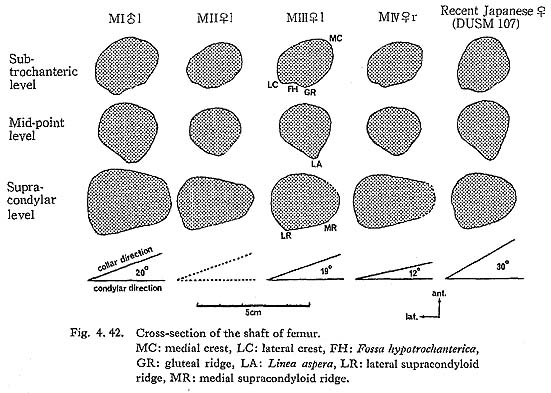
A marked gluteal ridge (Crista hypotrochanterica), (Figs. 4.40-C, 42) is seen in MIrl, the most proximal part of which forms the third trochanter, as described before, and the lower part of which continues to the lateral lip of the Linea aspera. In MIIl, MIII rl and MIVr this ridge is not remarkable. In MIIIrl a sharp Fossa hypotro-chanterica (, 1937) and swelled lateral gluteal flange (Angulus lat. sup., Klaatsch, 1901; Crista lateralia, Weidenreich, 1941) are seen (Figs. 4. 40-D, E, 42-FH, LC). Generally speaking, the appearance of the subtrochanteric region of MIIIrl is exactly the same as that of Sinathropus I according to the description by Weidenreich (1941). In MIrl, MIIl and MIVr the fossa and the flange are also seen to a lesser degree.
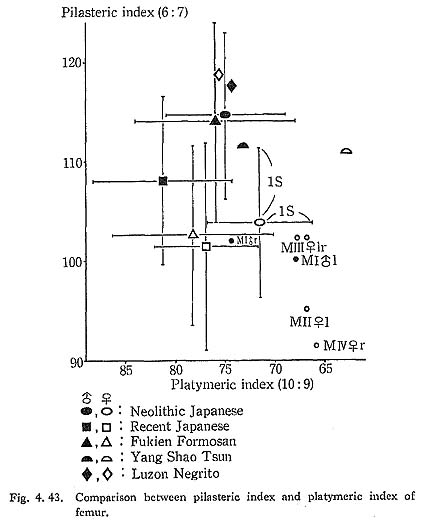
Because of development of the Crista medialis and lateralis, an outline of the cross-section becomes ellipical, the longer axis of which is in an antero-medial to postero-lateral direction (Fig. 4. 42). The platymeric index falls within the range of 71-73 except in MIr (79). These figures are far smaller than the means for the Neolithic and Recent Japanese or Fukien Formosan and Luzon Negrito, showing a considerable degree of platymery (Tables 4. 24, 25; Figs. 4. 42, 43), but they are larger than the means for the Yang Shao Tsun. In MIr this index becomes small because of the well-developed gluteal ridge.
The spiral line for the M vastus femoris medialis is seen in MIrl and MIIIrl, but is almost invisible in MIIl and MIVr. The pectineal line is not well developed in any of the specimens.
Mid-shaft
The femoral shaft in Minatogawa is, in general, cylindrical in the middle region, having a weak eminence at Linea aspera (Fig. 4. 42).
Generally speaking, as seen in most of the fossil Sapiens, the Linea aspera of the Neolithic Japanese is remarkably developed forming a pilaster which has two parallel lips (Crista aspera) for the vastus femoris. In the Recent Japanese, the same condition as in the Neolithic is seen to a lesser degree in matured males.
In MIrl the Linea aspera is considerably developed to provide double lips (Fig. 4.42). The distance between the two lips is 7 mm at the mid-shaft. The lateral lip, having a weak Fossa pilastrica, is slightly stronger than the medial one. In MIIIrl the devel-opment of these two lips is not remarkable, but there is a weak ridge between them. In MIIl and MIVr the linea is quite weak, showing only a low round eminence (Fig. 4. 42).
The pilasteric index of MIrl is far smaller than the means of the Neolithic and Recent Japanese males or other male populations (Table 4. 24, Fig. 4.43). The index of MIIIrl is close to the means of those female populations. In MIIl and MIVr, the indices are 95 and 91 respectively, which are quite small compared with those for the other female populations studied (Fig. 4. 43).
Roughly speaking, the Linea aspera in Minatogawa Man is very weak in spite of the well-developed trochanteric region. The appearance of the posterior surface of the shaft in female specimens (II, III, IV) would particularly indicate that they were quite young because of the very weak development of the Linea aspera and smoothness of its surface.
Supracondylar region
As described before, a slight widening in the lower shaft is seen in all the specimens. The degree of widening is relatively high in MI, II, VI and less in MIII (Fig. 4. 42). Above the condyle also, the shaft forms a gradual widening which is called "trumpet form" by Klaatsch (1901). This trumpet form is a characteristic of Recent Sapiens showing striking contrast to the sudden widening in the Classic Neanderthals.
The Fossa suprapatellaris is well-defined in MIrl and MIIIrl, but not as deep as in the Classic Neanderthals. No fossa is seen in MIVr. The upper part of the popliteal surface is flat or slightly concave in the present specimens (Fig. 4. 42). This character indicates also that the Minatogawa femora are close to the Recent Sapiens and apart from the Classic Neanderthals.
In the lower part of the popliteal surface, there is a low tubercle for the medial head of the gastrocunemius in all the specimens. The tubercle for the plantaris is not visible.
Lower Epiphysis
Intercondylar fossa
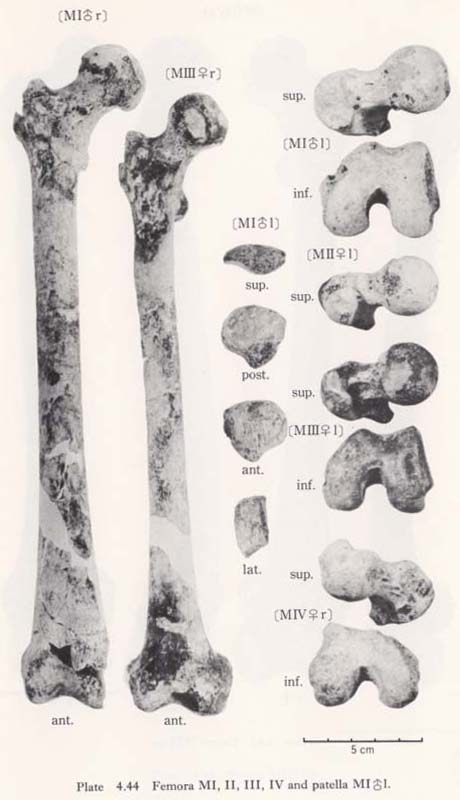
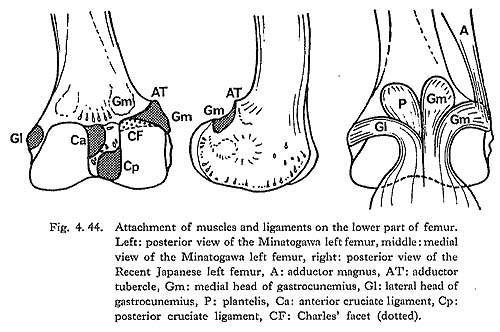
In cases of the lower ends of the Minatogawa femora, the most impressive feature is a narrowness of the intercondylar fossa (Fig. 4. 44). In usual cases the width of the intercondylar fossa is roughly equivalent to that of the medial condyle, while in the Minatogawa femora, the width of the former is far smaller than that of the latter. For example, 14 and 26 mm in MIl, 16 and 23 mm in MIIIr, respectively.
The biepicondylar width and the thickness of lateral condyle are 72 mm and 58 mm in MIl giving a condyle index of 81 between them, which is not very large as com-pared with the mean for the Neolithic Japanese males (75.9). In MIIIl these values are 65 mm and 54 mm giving an index of 83, which is close to the mean for the Neo-lithic Japanese females (79.5).
The narrowness of the fossa is, therefore, due not only to the small size of the lower epiphyseal width but also to the bulging of the condyle toward the fossa. In spite of the narrowness of the fossa, the facets which give rise to the cruciate ligaments are so wide that the knee joints of the Minatogawa people had to be robust (Fig. 4. 44).
Origin of the gastrocunemius muscle
In all the specimens there is a well-defined wide area which gives rise to the tendi-nous portion of the medial head of the gastrocunemius between the adductor tubercle and the medial condylar surface (Fig. 4. 44-Gm). Accordingly, it can be considered that the Minatogawa people had strong gastrocunemius muscles. This area is also seen in the Neolithic Japanese in a similar condition, but in the Recent Japanese it is less developed. The area for the origin of the lateral head is also seen on the lateral condyle as in usual cases of the Recent Japanese (Gl).
Charles' facet
There is sometimes an extension of the articular surface on the anterior end of the posterior surface of the medial condyle in populations of habitual squatters. This sur-face is called Charles' facet, after Charles (1894a), and has been considered to be formed by the compression of the medial meniscus in a flexed position of the knee joint, This facet is seen in all the Minatogawa femora (Fig. 4. 44-CF). The base length and the height of the facets are 11 and 8 mm in MIl, 11 and 6 mm in MIIIl, and 16 and 9 mm in MIVr.
Other characters
The trochlea for patella is quite deep and wide in MIrl and MIIIrl, which is similar to that of the Australian cited by Martin (1928). In MIIIl and MIVr, the condition is not clear because of damage. A marking by meniscus is relatively sharp in the anterior border of both condylar surfaces in the case of Minatogawa femora.
Internal Structure
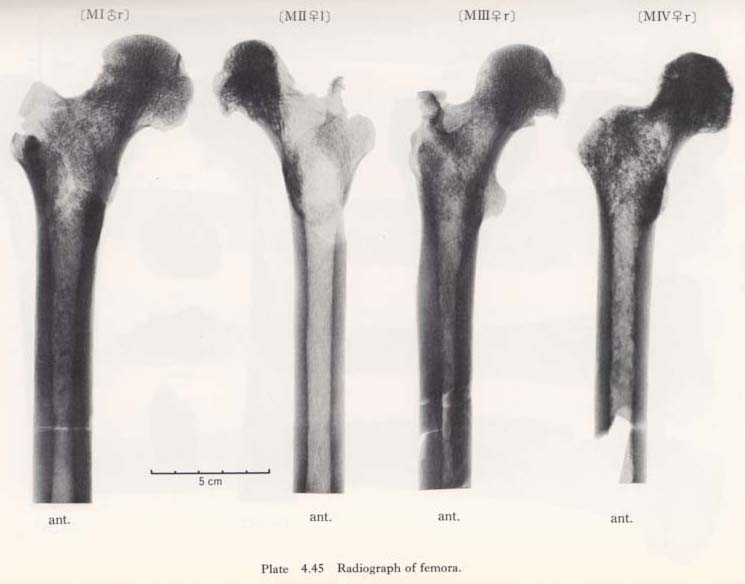
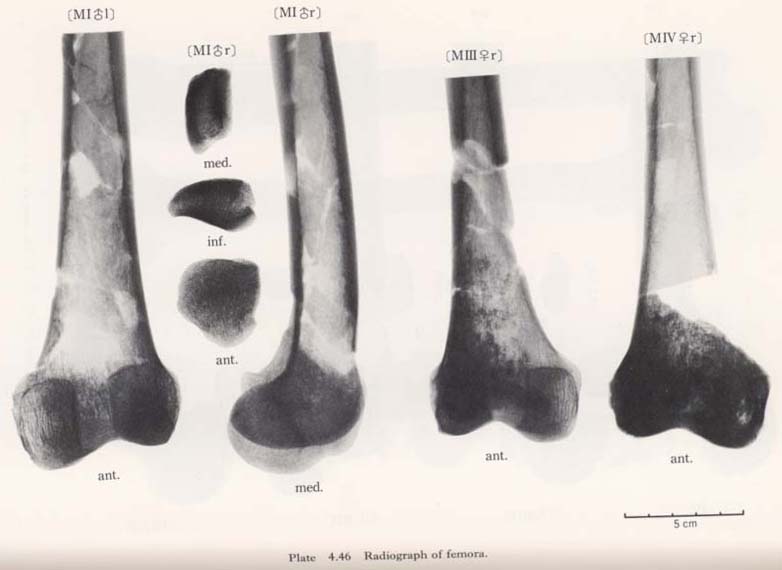
According to the sagittal radiogram of the Minatogawa femora, the so-called Ward's triangle is clearly observable, showing a typical character of the Recent Sapiens.
The thickening of the compact wall of the shaft is remarkable in MIIIrl, resembling the Sinanthropus femora according to the description by Weidenreich (1941) (Plates 4. 45, 46). In MIrl the thickening is also considerable and the thickest part is not in the mid-shaft but in the upper part of the shaft; whereas in MIIl and MIVr no thickening is seen.
Generally speaking, the trabeculae in these bones are bolder than in the Recent Japanese. However, it is likely that this boldness of the trabeculae does not correctly represent the condition while the individual is alive, but could be due to the fossil-ization, since the degree of density varies in each bone according to the portions.
Discussion
Constriction of the neck
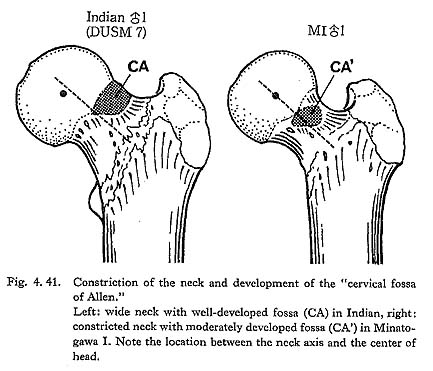
Generally, the constriction of the neck is caused mainly by the depression of the anterior and superior surface of the neck. Thus, the relative arrangement between the neck and the head seems to have changed by degree of this constriction.
In case of the constricted neck, the center of the head is located approximately on the extension of the neck axis when the bone is anteriorily seen (Fig. 4. 41). This type of arrangement is widely observable in such older fossils as Neanderthals as well as nonhuman primates. The constriction increases the range of movement on the hip joint, particularly in a flexed or abducted position. Accordingly, the range of the hip joint movement in the Minatogawa Man seems to be quite wide.
In case of the wide neck, the center of the head is located below the extension of the neck axis. This type is seen in the Recent Sapiens, especially in European whites, and it resembles the condition of cursorial mammals as carnivor. It seems that the wide neck is adapted mainly for the bending moment acted on the neck along the frontal plane at the expense of movement range.
Accordingly, it is natural that the constricted neck is not strong for the bending moment acted on the neck by the body weight and muscle contraction. But it does not directly indicate that those who have the constricted neck have a light body weight or weak muscles. For example, the constricted neck often combined with the robust shaft as in the Neanderthals and the Neolithic Japanese as well as great apes. In these cases, it should be considered that the bending movement acting on the neck is canceled or lightened by the reverse bending moment caused by the hip muscles as the piriformis, gemerus, obturators and femoral quadrate.
Cervical fossa of Allen
The cause of the formation of the cervical fossa of Allen is considered as the com-pression of the antero-superior surface of the neck by the acetabular brim in a flexed position like squatting. Thus, as far as the occurrence and duration of keeping such position is equal, the development of the fossa becomes high, being directly propor-tional to the width of the neck, or to the degree of elevation of this region (Fig. 4. 41). Actually, no fossa is seen on the fully constricted neck of nonhuman primates in spite of their high degree of habitual squatting.
On the contrary, the fossa is often remarkably developed on the wide neck of the Neolithic and Recent Japanese, which gives the sign of habitual squatting or square sitting on tatami (Japanese mats). In Minatogawa femora, the development of the fossa is moderate, but making allowance for the constriction of the neck, this may suggest a considerable degree of habitual flexion of the hip joint like squatting.
Platymery
As noted already, the subtrochanteric cross-section is elliptical giving the platymeric indices from 71 to 73 except in MIr (79). Its long axis is in the same direction as the neck axis in all the Minatogawa femora. It seems to us that this flatness (platymery) is a biomechanical adaptation to the bending moment caused by the complex force acted on the head and trochanter region.
Linea aspera
Generally speaking, there is a certain tendency for the Linea aspera to develop proportionally to the increase of the attaching muscles (adductors, femoral vasti and short head of biceps) in the case of the modern Sapiens. In accordance with this proportion, it is likely that these muscles in Minatogawa people may be weak, especially in MIIl and MIVr. However, at least the rectus femoris and long head of the biceps femoris could be well developed, because the inferior anterior iliac spine and the ischial tuberosity are considerably developed even in MIIl.
There may be two ways of interpreting this situation in the Minatogawa femora. First, these attaching muscles might be fairly developed in spite of poor development of the Linea aspera. Second, they might not be as developed as the rectus femoris and long head of the biceps.
Judging from first interpretation, there seems to be a certain difference in the biological mechanism to develop Linea aspera between modern Sapiens and such other older fossils as Sinanthropus, Neanderthals and Minatogawa man, yet the reason for such difference is unknown. From a second interpretation, it is rather difficult to explain why only a small part of the muscle group is disproportionate to the other. One answer may be as follows. The muscles which attach to the Linea aspera are single-joint muscles, whereas the rectus femoris and long head of the biceps are two-joint muscles; thus it seems possible that there is another way of adaptation in locomotion in which the two-joint muscles become more dominantly developed than the single-joint muscles.
Formation of Charles' facet
According to the studies on three cadavers, the tendon of the medial head of the gastrocunemius muscle arises from the area between the adductor tubercle and the margin of the medial condylar surface. The tendon runs medially, covering the posterosuperior part of the medial condylar surface to the intercondylar fossa, then turns inferiorly (Fig. 4. 44). This tendon and Charles' facet are located facing each other with a capsule between them. We have drawn the tentative conclusion about the formation of this facet that it is caused not by the compression of the medial meniscus but by that of the tendon of the medial head of the gastrocunemius in a flexed position of the knee joint such as occurs in squatting or other sitting postures.
Overall similarity to the other groups
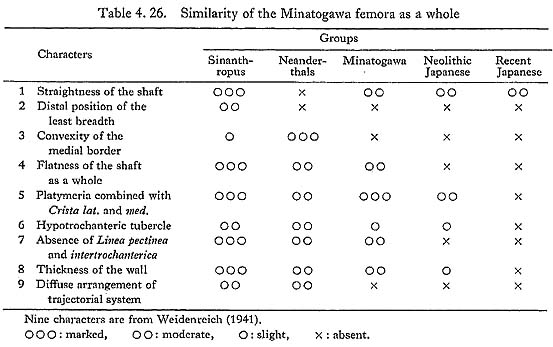
Weidenreich discussed (1941) the overall similarity of femora between Sinanthropus and modern man. He pointed out nine items which characterize the Sinanthropus femora as follows:
(1) straightness of the shaft
(2) distal position of the least breadth of the shaft
(3) convexity of the medial border of the shaft
(4) pronounced flatness of the shaft as a whole
(5) pronounced platymeria of the substrochanteric region, combined with angularity of the medial and lateral borders
(6) development of a hypotrochanteric tubercle at the base of the greater trochanter and its direct continuation into the hypotrochanteric crest
(7) absence of the Linea intertrochanterica and Linea pectinea
(8) extraordinary thickness of the wall of the shaft and neck, combined with a narrow calibre of the medullary canal
(9) diffuse arrangement of the trajectorial systems of the upper end combined with a tight meshed and coarse cancellous substance.
He pointed out that Sinanthropus differs from modern Sapiens in the above respects. He also stated that Neanderthals more or less resemble Sinanthropus in several traits (Nos. 6, 7, 8), but the former is completely apart from the latter in stoutness and pronounced curvature. He maintained "... Nor could the Neanderthal form be regarded as intermediate between Sinanthropus and modern man."
In the same way we try to compare the Minatogawa femora to Sinanthropus, Neanderthals, Neolithic and Recent Japanese using Weidenreich's nine characters. As in Table 4. 26, Minatogawa femora seem more or less similar to those of the Sinanthropus and Neanderthals, and also to some extent similar to those of the Neolithic Japanese. In characteristics Nos. 4, 5, 6, 7, Minatogawa man, Sinanthropus and Neanderthals have the same tendency and show apparent primitiveness. In Nos. 3 and 9, Sinanthropus and Neanderthal show a similar tendency. But the convexity of the medial border (No. 3) is very slight in Sinanthropus. The trajectorial system and cancellous substance (No. 9) do not always correctly represent the condition while the individual is alive, as already noted; thus, these two characters do not seem to be important.
Minatogawa femora show some similarity to Sinanthropus femora in diaphyseal characters, namely, straightness combined with distal position of the vertex of curvature and relative slenderness in spite of the thickness of the compact wall. Furthermore, they show marked difference from Neanderthals in straightness and slenderness. As far as the poor specimens in other Japanese Paleolithic skeletons are concerned, they show relative slenderness having rather short stature (Suzuki, 1962, 1966; Ogata, 1973), which is not contradictory to the characters of the Minatogawa femora. It may be said that the Minatogawa femora represent a certain stage of older Mongoloid femora which show more or less transitional features from Sinanthropus to Modern Mongoloid as Japanese, although of course they are closer to the latter.