CHAPTER 4
Postcranial Skeleton of the Minatogawa Man
Hisao Baba* and Banri Endo**
*Department of Anatomy, Dokkyo University School of Medicine;
**Department of Anthropology, Faculty of Science,
The University of Tokyo
| ( 5 / 7 ) |
|


  |
  |

Proportion
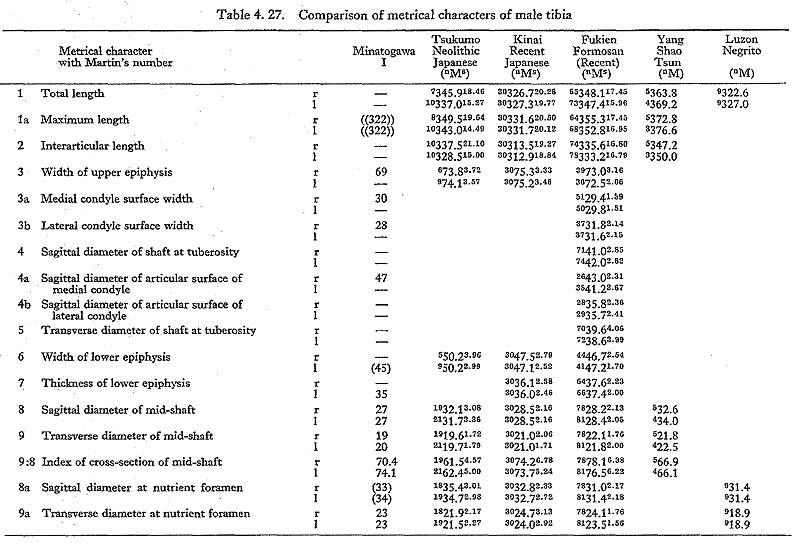
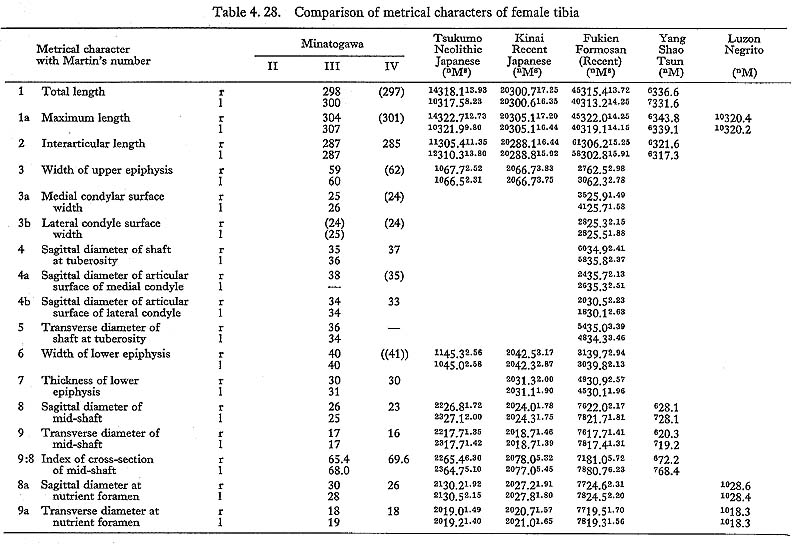
The total length of MI& was also estimated as 327 mm. The mean value of the least circumference in both sides is 70,5 mm. Accordingly, the length-thickness index is 22, being similar to the means for the Neolithic and Recent Japanese or Fukien For mosan males but rather larger than the mean for the Negrito. The index in both MIII rl and MIVl is approximately equal to the means for the females of those populations except Negrito, which has a lower mean.
rl and MIVl is approximately equal to the means for the females of those populations except Negrito, which has a lower mean.
Torsion angle of the shaft
This angle of MI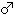 is roughly estimated as 19°This figure is roughly similar to the means for the Neolithic and Recent Japanese males, taking their widely varying range into consideration. It is also the case with MIVr. In MIII, this measurement varies in right and left bones (4° and 21°,respectively).
is roughly estimated as 19°This figure is roughly similar to the means for the Neolithic and Recent Japanese males, taking their widely varying range into consideration. It is also the case with MIVr. In MIII, this measurement varies in right and left bones (4° and 21°,respectively).
Upper Epiphysis
Size
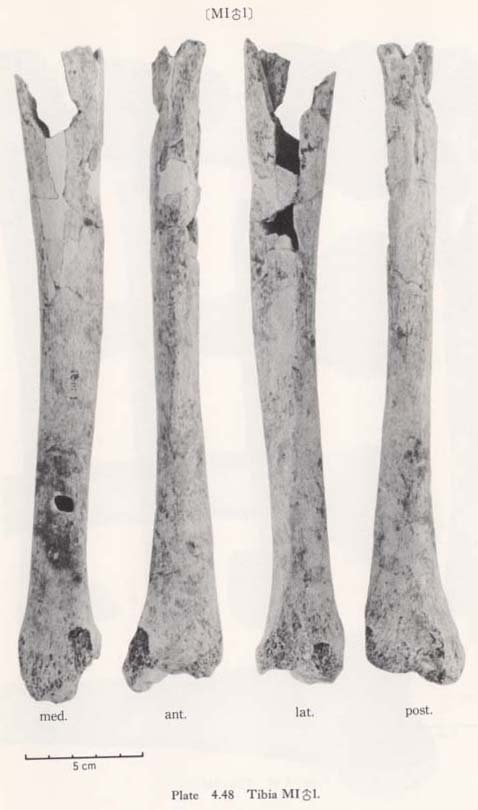
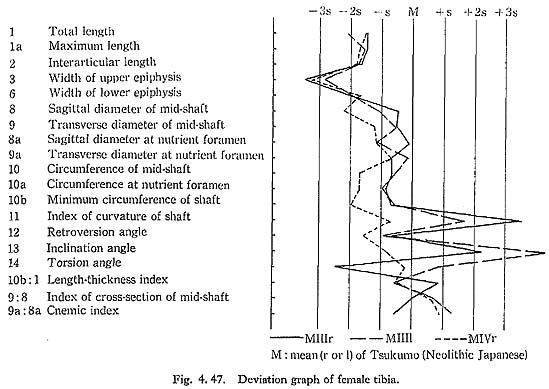
The upper epiphyseal width is generally quite small as compared not only with the means for the Neolithic and Recent Japanese or Fukien Formosan but also with its own length (Fig. 4. 48).
Position of the upper end to the shaft
The retroversion angle of the upper end of Minatogawa females is roughly equivalent to the means of Neolithic and Recent Japanese or Recent Fukien Formosan females. This indicates that the inclination (not retroversion) of the medial articular surface to the shaft is moderate. The biaxial angle (the difference between the retroversion angle and the inclination angle) indicates the real retroversion (backward displacement) of the upper end to the shaft. It is 3° in MIIIr, 4° in MIIIl and 3° in MIVr, being slightly less than the means for the females of those populations (Tables 4. 27, 28).
Articular surface
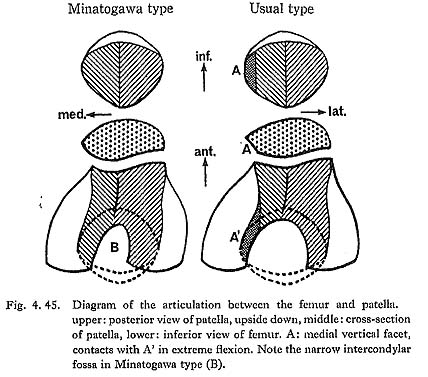
Both medial and lateral articular surfaces are wide in relation to the upper epiphyseal width. According to Thomson (1889), the lateral articular surface in a habitual squatter shows a marked convexity in sagittal section and a downward curvature on the posterior part of its articular surface. However, no convexity is seen in MIr or MIIIrl, although it is slightly convex in MIVr. MIIIrl and MIr show a moderate curvature on the posterior part of the articular surface. In MIr, the condition is not clear because of damage in this part.
The fibular facet in MIIIrl is rather small in size, and moderate in MIVr. In MIr the facet is broken off. The inclination angle of this facet from the horizontal plane is roughly 40° in MIIIrl and MIVr.
Intercondylar area
In relation to the structure in the lower end of the femur, the intercondylar area of the tibia is generally narrow in Minatogawa specimens. In order to show the width of the intercondylar area, the distance between the tops of both sides of the intercondylar eminence is useful, but no such comparative data have been published. Therefore, we estimated the intercondylar width by subtracting the sum of medial and lateral articular width from the upper epiphyseal width. As Table 4. 29 shows, the inter condylar width in Minatogawa tibiae is far smaller than the values for the Recent Kanto Japanese except in MIVr. There are sharp pits for the insertion of the menisci along the antero-medial margin of the lateral articular surface (for lateral miniscus) and the postero-lateral margin of the medial articular surface (for medial meniscus). These pits are often seen in the Neolithic Japanese but seldom seen in Recent Japanese.

Tuberositus tibiae
The tuberosity is well developed forming a prominent tuber in MIIIrl and MIV. In MIrl, it is not observable because of breakage. Its width is 18 mm in MIIIl, 21 mm in MIIIr showing relatively large values in spite of their slender shafts. Judging from the development of the tuberosity, it is likely that the quadricepus femoris muscle was relatively strong.
Other characters
The groove for the semimembranosus muscle is seen as in usual cases. It runs transversely on the postero-medial surface of the upper end in MIIIrl and MIVr. The area corresponding to the bursa for the popliteus muscle is not identified, because the depression under the postero-medial margin of the lateral articular surface is situated so high that it almost contacts the articular margin in all the Minatogawa specimens.
Diaphysis
Form as a whole
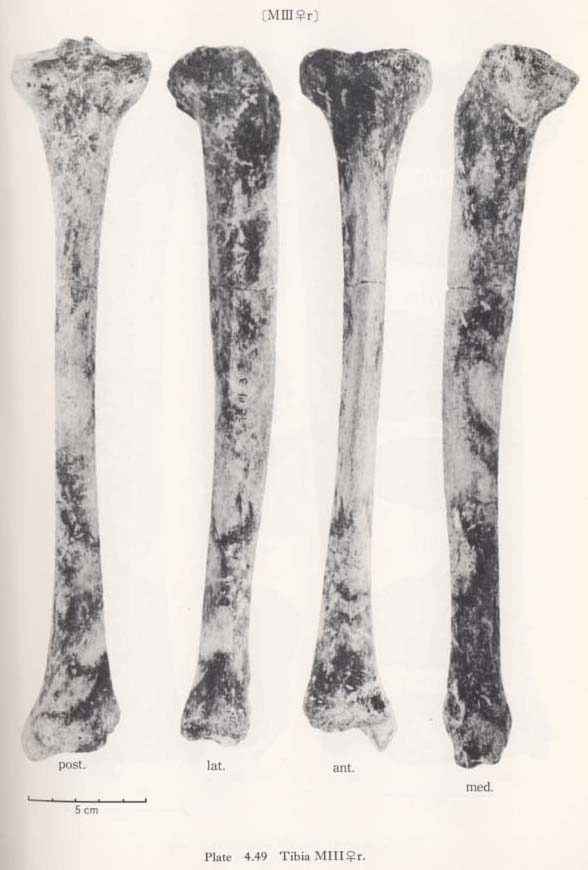
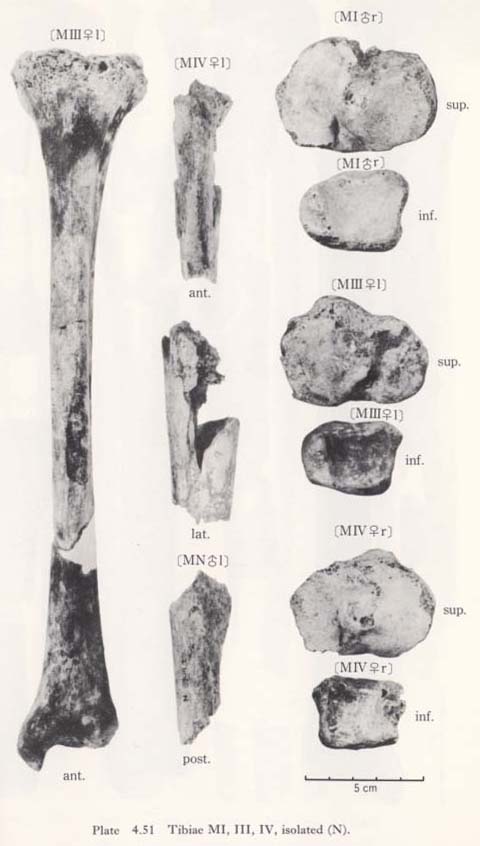
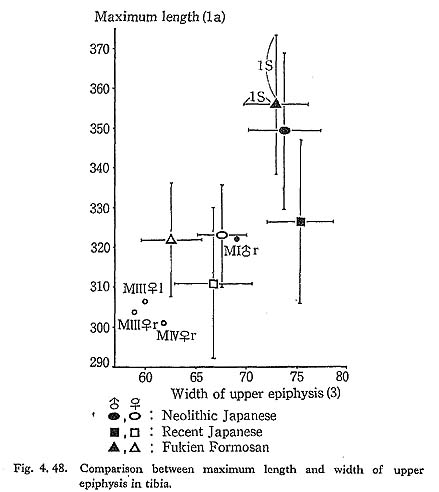
There is a peculiar feature in the shafts of Minatogawa tibiae in their front-view, namely, the medial convexity (Fig. 4. 49). In usual cases, the lateral and medial borders of the shaft are straight or slightly concave, while the lateral border is quite concave and the medial one is convex in this case. Accordingly, the median line of the shaft is medially convex in Minatogawa specimens. Such a pronounced convexity cannot be seen in the Neolithic and Recent Japanese tibiae stored in the University Museum of the University of Tokyo except in a few cases which show a slight medial convexity. There are some reports about this medial convexity in the Classic Neanderthals such as La Chapelle-aux-Saints and La Ferrassie I. As for the antero-posterior curvature of the shaft, the Minatogawa specimens show a considerable variability. In MIIIrl, the curvature index is 3.1, and these have a sharp anterior flexion at the mid-point. This figure is fairly large as compared with the means for the Neolithic and Recent Japanese or Fukien Formosan females. In MI, estimated value is roughly 2 and is moderate as compared with that of the Neolithic and Recent Japanese males or Recent Fukien Formosan.
Cross-section
The sagittal diameter at the mid-shaft of MIrl is not only far less than the mean for the Neolithic Japanese but also slightly less than that of the Recent Japanese and Fukien Formosan or Yang Shao Tsun males, while the values of MIIIrl are slightly smaller than the means for the Neolithic Japanese and Neolithic Yang Shao Tsun females but larger than those for the Recent Japanese and Fukien Formosan females.
The transverse diameters of the mid-shaft of Minatogawa specimens are rather close to the means for the populations compared.
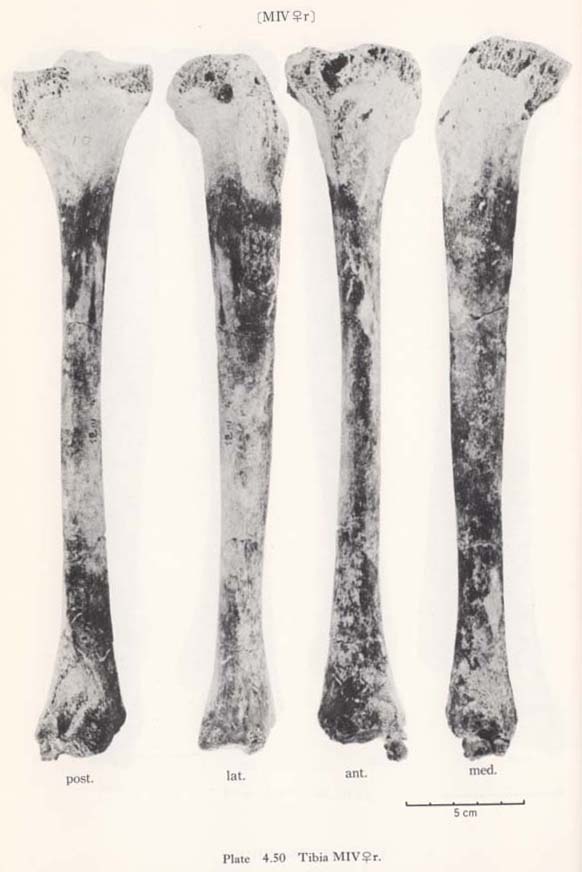
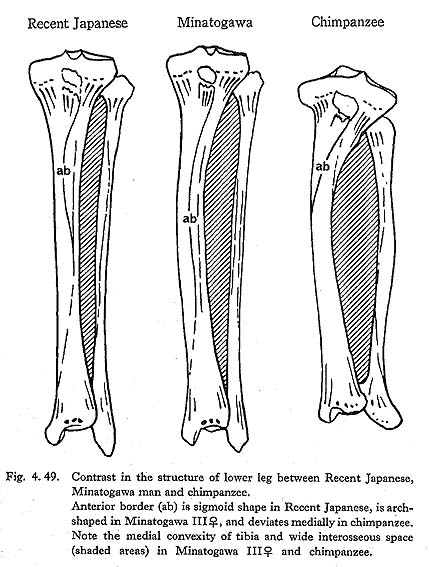
The cross-section index of mid-shaft in MIr is larger than the means for the Neolithic Japanese or Yang Shao Tsun males and rather close to those of the Recent Jap anese and Fukien Formosan males (Fig. 4. 50). This index in MIIIrl and MIVr is roughly comparable to the mean for the Neolithic females and far less than those for the Recent Japanese and Fukien Formosan females.
The sagittal diameter at the level of the nutrient foramen in MI falls between the means of the Neolithic and Recent Japanese males. The same is true of MIII. Only the values of MIVrl are rather close to those for the two Recent female populations.
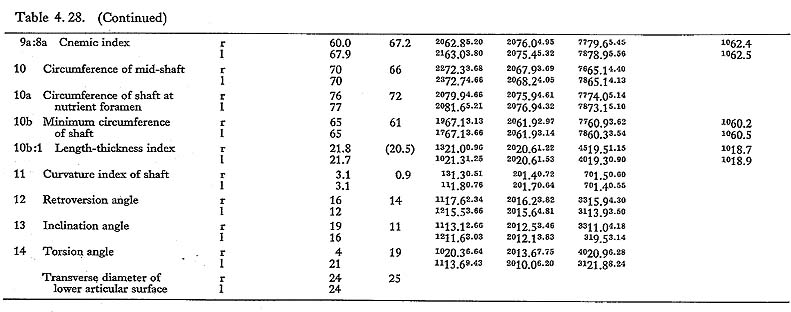
The cnemic index at the nutrient foramen (cf. Manouvrier, 1887) varies from 67 to 69 except for MIIIr (60). This range is generally comparable to the means for the Neolithic Japanese males and females, and smaller than those for the Recent ones (Fig. 4. 50).
In general, the Minatogawa tibiae show a considerable degree of platycnemia in comparison with the Recent Japanese but fall within the range for the Neolithic Japanese.
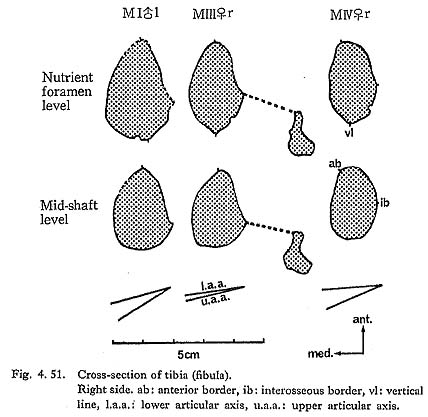
The contour of the cross-section at the level of the nutrient foramen is somewhat rhombic combined with the vertical line which will be described later (Fig. 4. 51). The contour of the mid-shaft is not rhombic.
Posterior surfaces
The soleal line is marked in MIr and MIVr forming a narrow but sharp ridge, while it is rather poor in MIIIrl. In these cases, the vertical line for apponeurosis covering the tibialis posterior runs from the mid-point of the soleal line downward along the median line then turns laterally and, after passing the mid-point of the shaft, disappears near the interosseus margin.
Lateral surface
In MIrl and MIIIrl, the interosseus border is sharp. Anterior to it, there is a wide vertical fovea on the lateral surface of the tibia in MIIIrl. and MIIIrl. Since the interosseus border is straight, the concave posterior border comes close to this border at mid-shaft level in MIrl and MIIIrl. In MIVr, the border is weak and situated along the straight median line of the shaft.
Anterior border
In usual cases, only the upper part of the anterior border for the apponeurosis of the tibialis anterior is situated medially across the center of the anterior bony mass of the shaft forming a sharp edge (Fig. 4. 51). The border, therefore, shows a sigmoid line in shape from an anterior view (Fig. 4. 49). In MI and MIII the anterior border is not situated as much across the anterior bony mass, because the shaft itself situates medially by its medial convexity. In MIVr, the border comes across the anterior bony mass not only in the upper shaft but also in the lower shaft, presumably being combined with the narrow space for the extensor muscles by the anterior position of the interosseus margin and the poor development of the fovea on the lateral surface.
This fact suggests that the position of the extensor muscles is situated relatively low in comparison with the Neolithic and Recent Japanese. This type of anterior border is seen in Classic Neanderthals such as Spy, La Chappelle-aux-Saints, La Ferrassie (Boule, 1911-13). The anterior border is sharp in MI rl and blunt in MIII rl and MIVr.
Lower Epiphysis
Size
The maximum width of the distal end in the Minatogawa specimens is far less than the means for the Neolithic Japanese males and females and slightly less than those for the Recent Japanese. These values are rather close to that for the Fukien Formosan.
Squattmg facets
Articular facets or impressions are often seen in the antero-interior part of the tibia along the anterior margin of the inferior articular surface in a habitual squatter (Thomson, 1889; Barnett, 1954; Morimoto, 1959; Baba, 1970). Since these areas act as stoppers by contacting the corresponding areas on the talus in extreme dorsiflexion of the ankle joint as in squatting, they were designated, together with the areas on the talus, as squatting facets.
The facets in Japanese can be divided into four kinds (Fig. 4. 52):
A. lateral (squatting) facet: A facet situated in the inferior fossa described later. It contacts the lateral part of the talar neck (lateral facet of the talus-A').
B. medial (squatting) facet: A facet situated in the antero-medial corner of the inferior articular surface. It contacts the anterior extension of the medial part of the trochlear surface of the talus (medial facet of the talus-B').
C. medial malleolar (squatting) facet: Medially curved anterior part of the medial malleolar surface. It contacts the anterior part of the medial malleolar surface of the talus (medial malleolar facet of the talus-C').
D. inferior fossa: Not an articular surface but impression situated anteriorly and superiorly to the anterior margin of the inferior articular surface. It contacts the superior surface of the talar neck (Processus trochlearis-D').
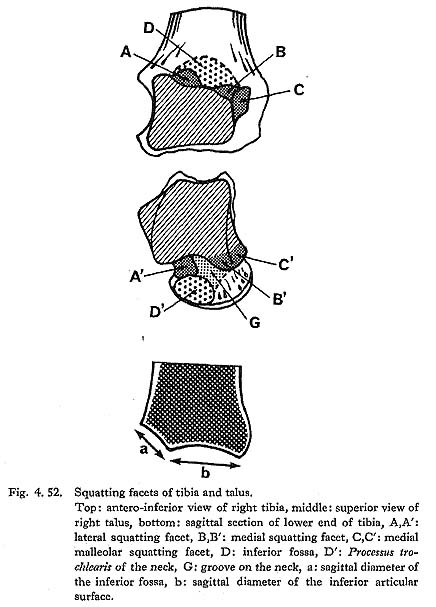
A marked lateral facet is seen in MIl, expanding the whole area of the inferior fossa. It is also seen in the lateral part of the inferior fossa in MIII.rl and MIr. The medial facet is not seen in MIl, while it is slightly developed in MIIIrl. The condition in MIVr is not clear because of breakage. The medial malleolar facet is not distinct in MIl and MIIIl and is very poorly developed in MIIIr. The corresponding part of MIVr is broken. The inferior fossa is seen in all the specimens as moderately developed. Generally speaking, the development of the squatting facet is not as pronounced as in the Neolithic Japanese and is approximately equal to that in the Recent Japanese.
Other characteristics
The surface of the Incisura fibularis in the Recent and Neolithic Japanese is sometimes inclined infero-laterally, but is vertically situated in the Minatogawa specimens. There are two tubercles for the tibio-fibular ligments which form an anterior and posterior margin of this notch. These tubercles in Minatogawa specimens are not as prominent as in the Recent and Neolithic Japanese.
The groove for the tibialis posterior tendon is distinct but not deep in these tibiae. Its surface is roughened, presumably by the absorption and hyperplasia of bone. The groove for the tendon of the flexor hallicus longus is seen only in MIVr.
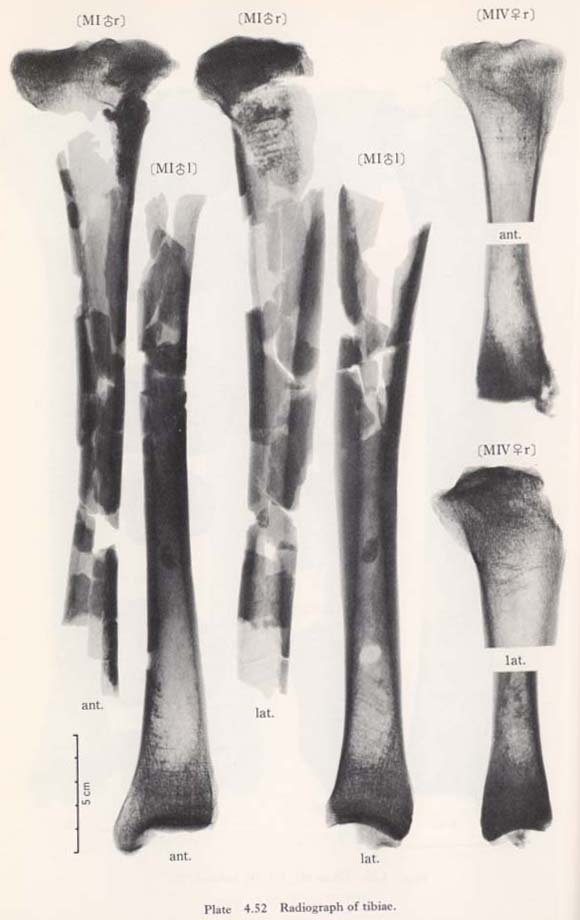
Internal Structure
As shown in the transverse and sagittal radiogram, the compact wall is extraordinarily thick in MIIIrl, while in MIrl and MIVl, the wall is not very thick. Several numbers of the so-called transverse line (Harris's lines) are seen in both upper and lower parts of the shaft in MIrl and MIIIrl. They are also seen in MIVr to a lesser degree.
MNl fragment
This fragment had first been classified as MIII. It occupies the postero-lateral part of the upper shaft; its height is 72 mm and the width is 28 mm. An interosseus margin, a soleal line and a nutrient foramen are observable.
The interosseus margin is well defined but not prominent. The soleal line is well developed forming a wide ridge. The diameter of the nutrient foramen is slightly larger than that of MIl. The thickness of the compact wall is approximately equal to that of MIrl, From these facts, it seems that this fragment belongs not to a female specimen (MII) but to another unknown male specimen.
Discussion
Medial convexity of the shaft
The medial convexity serves to widen the interosseus space between the tibia and fibula (Fig. 4. 49). As for the anterior part of its space, the wide fovea on the lateral surface also serves to widen the space. Accordingly, it is probable that the extensor muscles such as the tibialis anterior, extensor hullucis longus and extensor digitorim longus which occupy this space were well developed in Minatogawa people.
In the posterior part, it also seems that the flexor muscles such as the tibialis posterior and flexor digitorum longus which are situated in the posterior part of this space may also be strong. This inference may be reinforced by the presence of a vertical line. The medial convexity is seldom seen in modern Sapiens but is sometimes seen in Neanderthals. In addition, it is quite common in nonhuman primates.
Anterior convexity of the shaft
There are two interpretations of the anterior convexity of the tibial shaft. The first is that it is related to the squatting posture. If the anterior convexity of tibia and femur is strong, the pressure caused by the flexor muscles of thigh and leg in extreme flexion of the knee joint such as in squatting is weaker than in the case of a straight tibia and femur. Thus, the anterior convexity is regarded as an advantageous character for a habitual squatter, particularly for those with well-developed flexor muscles.
In spite of the fact that the thigh and leg muscles are relatively thin and the anterior convexity of the femur is pronounced, the tibia is straight. Actually, in Indians who have thin muscles, markedly developed squatting facets often coexist with the convex femur and straight tibia. Reversely, when the femur is straight and the muscles are thick, the tibia seems to be convex.
This tendency may be applicable to the present specimens. MI and MIII have straight femora, convex tibae and relatively well-developed muscles, while MIV has a relatively convex femur, straight tibia and poor muscles.
The second interpretation is based on the function of the flexor muscles in the leg. The flexor muscles inserting the posterior surface of the mid-shaft as the tibialis pos terior and flexor digitolum longus act to make the shaft posteriorly convex by pulling the mid-shaft infero-posteriorly, while the body weight and other muscles such as gastrocnemius act to make the shaft convex anteriorly. Thus, if the flexor muscles inserting the mid-shaft are relatively strong, the shaft will become anteriorly convex. In MI and MIII, these muscles are considered to be strong because of the wide interosseus space.
Inferior fossa and the squatting index
The general development of the squatting facets in the Minatogawa specimens has already been described, but the development of each facet is so variable that estimation of the degree of habitual squatting is rather difficult. Since the inferior fossa is made by the compression of the superior surface of the talar neck, it serves not only to act as a stopper but also to widen the rotation angle of the ankle joint in extreme dorsiflexion. Accordingly, those who have well-developed inferior fossa can maintain the squatting posture easily for a long time.
Therefore Baba has attempted to give a "squatting index," a figure which represents the degree of development of the inferior fossa (Baba, 1970). This index is given by the ratio of the sagittal length of the inferior fossa (Fig, 4. 52-a) to that of the inferior articular surface (b). As shown in Table 4. 30, the squatting index for Minatogawa specimens is located between the means fo'r the Neolithic and those for Recent Japanese, which indicates that the degree of habitual squatting is intermediate of these two groups.

The transverse (Harris's) lines
According to the sagittal radiograph, there are more than seven lines running hori zontally in the upper and lower shaft of all specimens, except that the lower shaft of MIVr is in poor condition. The number of lines are far more than those in European groups listed by Wells (1967) and also those in Japanese by Hojo (1976).
By the observation of the transverse radiograph of MIrl and MIIrl, these lines are not horizontal as in sagittal radiograph but bear much resemblance to the (pattern of) trajectorial lines. In the transverse radiograph of the upper shaft of MIVr, they are rather horizontal, showing no similarity to the trajectorial lines (Plate 4. 52). These lines are considered to be formed by repeated illness or episodes of starvation (according to Wells, 1967). But as far as the present specimens are concerned, the cause of the formation of these lines is not clear.