CHAPTER 4
Postcranial Skeleton of the Minatogawa Man
Hisao Baba* and Banri Endo**
*Department of Anatomy, Dokkyo University School of Medicine;
**Department of Anthropology, Faculty of Science,
The University of Tokyo
| ( 1 / 7 ) |
|







    |
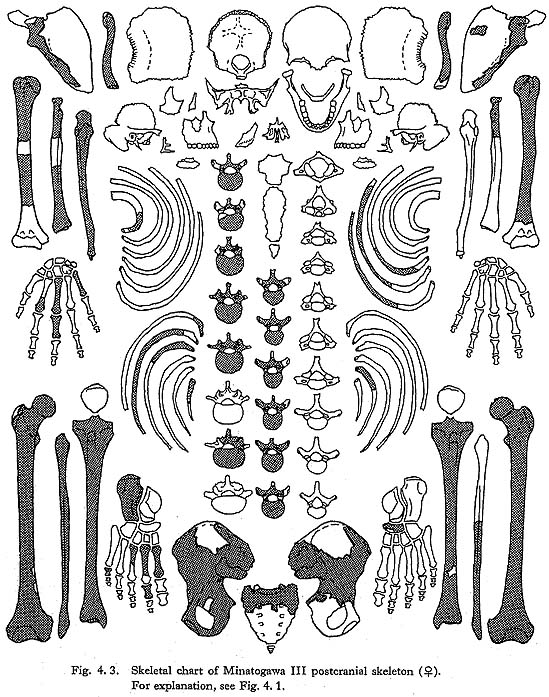
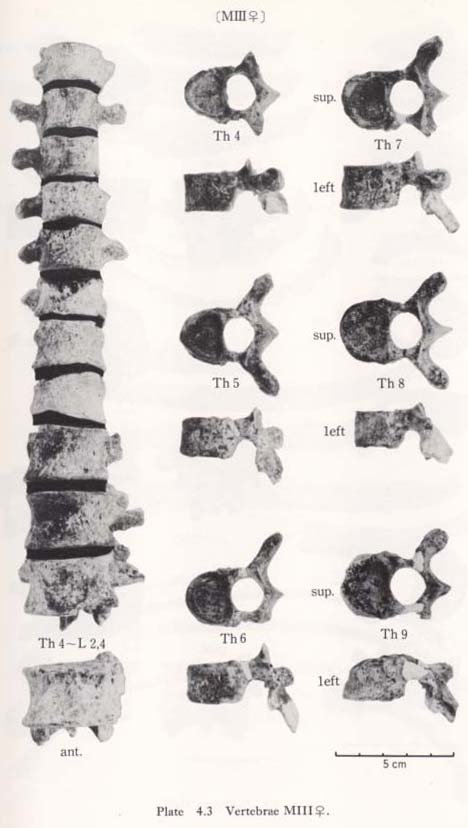
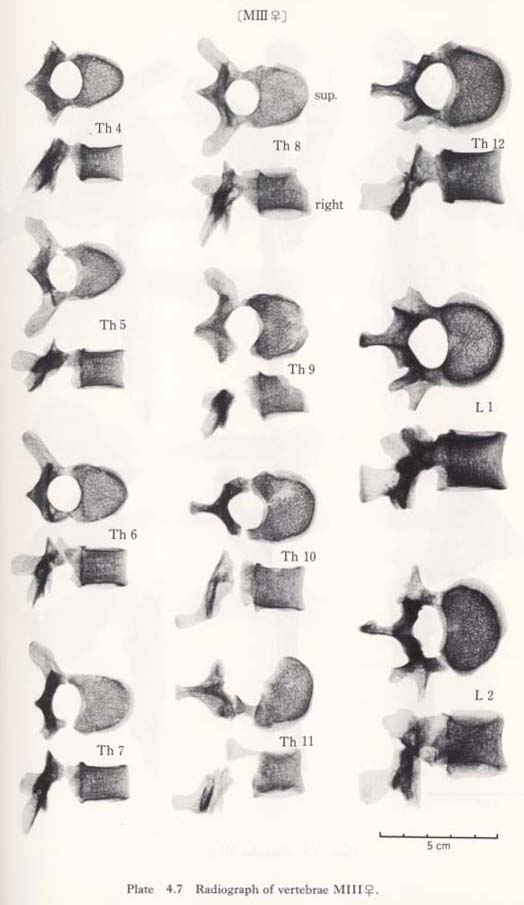
Fourth to twelfth of MIII 
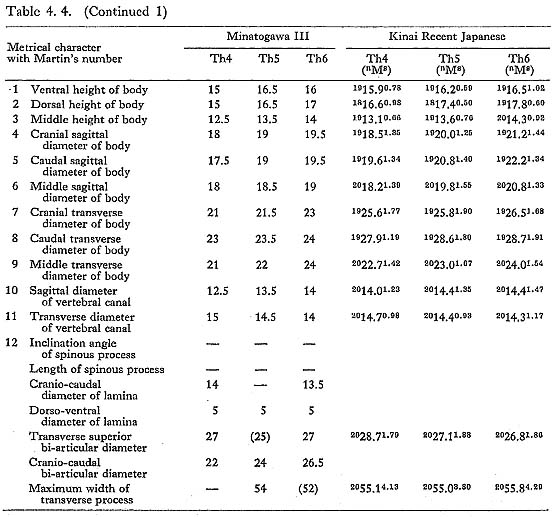
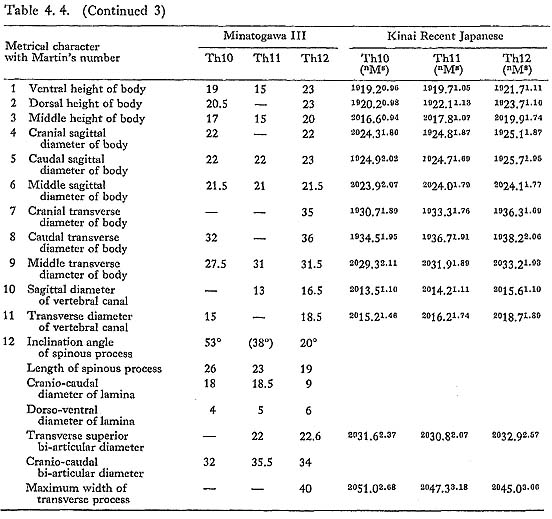
These bones are relatively well preserved and bear a generally close resemblance in character to those of the Recent Japanese females. Their sizes are slightly smaller (Table 4. 4). A peculiar feature is seen on the body of the eleventh bone: the right anterior parts of the upper and lower surfaces are depressed, forming sharp pits (Fig. 4. 6). The corresponding parts of the lower surface of the tenth bone and the upper surface of the twelfth are swollen to fit with the pits. This may suggest that the depression did not occur as the result of an injury.

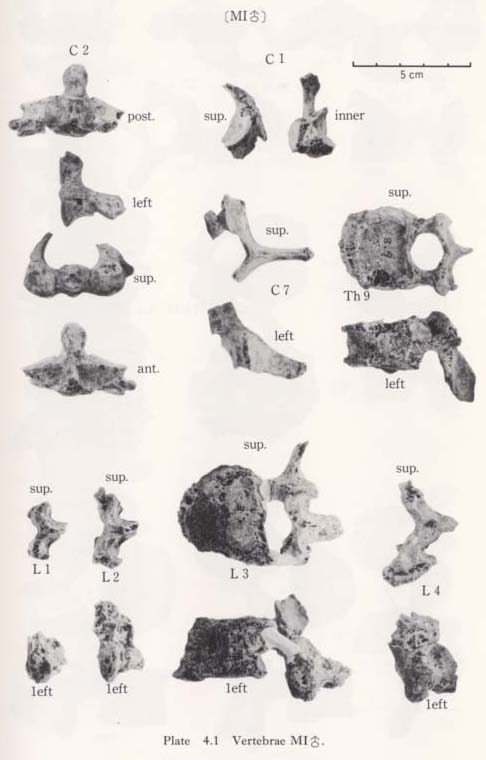
Ninth (?) thoracic vertebra of MI
This bone seems rather stout, but there are many changes, probably due to aging. There are lippings of osteophytes along the margin of the upper and lower surfaces. The body is compressed and numerous small hollows are seen on the articular facets showing light osteoporosis. The size of the body is similar to that of the ninth thoracic in the Recent Japanese males. Its facets for the costae are circular on the left side and hemicircular on the right. The spinous precess inclines strongly downward like those of the middle thoracic vertebrae, but it is not as long as the middle thoracics (Table 4. 3). From these facts, this bone may belong to the ninth thoracic vertebra.

Twelfth thoracic vertebra of MII
The remaining part of this bone is a small fragment of the body. It will be described later together with the series of the, lumbar vertebrae from MII.
Lumbar Vertebrae
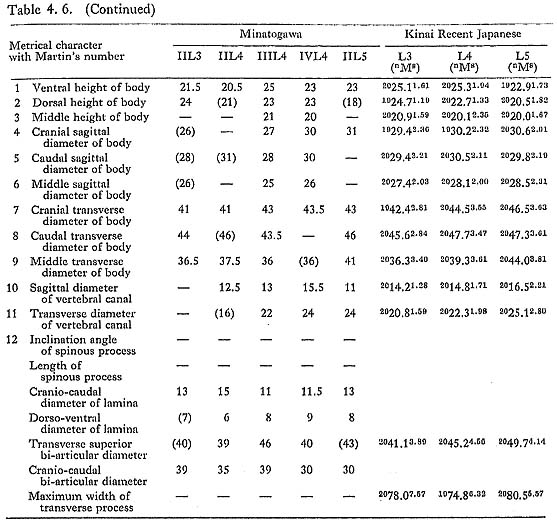
We have a whole series of lumbar vertebrae from MII. These bones were tightly cemented by carbonate of limestone with small flakes of the twelfth thoracic and the sacrum. In addition, there are the first, second and fourth from MIII in good condition, the first to the fourth from MI in poor condition, and the second and the fourth from MIV in rather good condition.
First and second lumbar vertebrae
The sizes of the preserved female bones are roughly equivalent to those of the Recent Japanese females (Table 4. 6). The anterior part of the body of the first lumbar vertebra from MII is strongly compressed. From a lateral view, the body appears wedgeshaped. This deformation may be due to the osteoporosis, because the lippings of osteophytes and the degenerative change are marked in this specimen. The remains of MI are only parts of the arches from the first and second bones. Numerous small hollows are seen on their articular facets. The sex of the second lumbar identified as being MIV. is actually not clear. Its overall size is small but the pedicle is thick (10 mm) and the crest between the upper and lower articular processes is very strong, indicating a male character.
  |
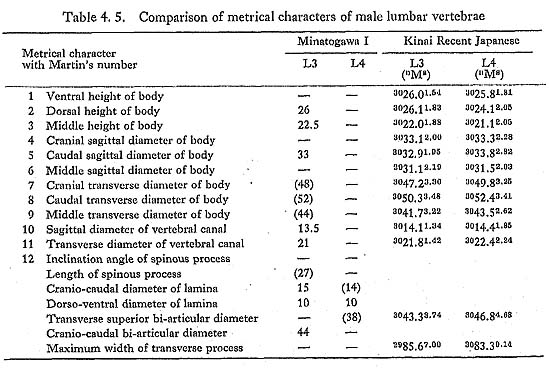
Third to fifth lumbar vertebrae
All the female specimens are almost the same size or slightly smaller than the Recent Japanese (Table 4. 6). The third bone from MI is large in size and seems generally stout (Table 4. 5). The lippings of osteophytea and the hollows on the articular facets are fairly developed. These lippings are also visible on the third, fourth and fifth vertebrae from MII.
Vertebral Column
Height of the vertebral column
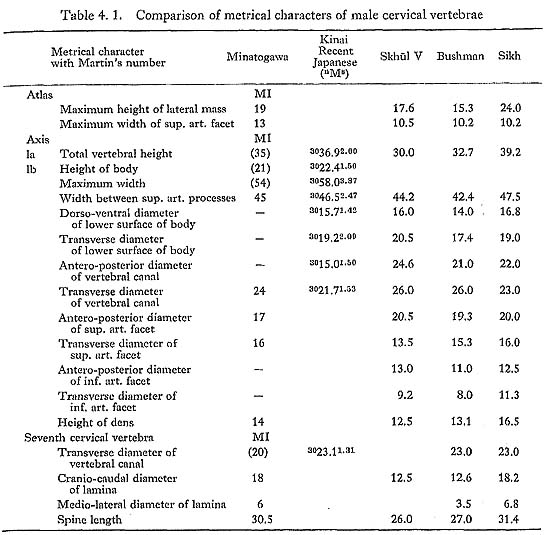
Our materials for estimating column height are poor. For the thoracic region, only MIII has provided us with data. In this case, the ventral body height can be measured from the bones of the third to the twelfth, except the ninth. The sum of the above heights is 137 mm, which is 96% of the corresponding sum of the means for the Recent Japanese females (Table 4. 6).
For the lumbar region, two individuals are available. The sum of the ventral body heights in the second to the fifth from MII is 87 mm. This value is 89% of the corresponding sum for Japanese females. The sum of the same heights of the first, the second and the fourth lumbars from MIII is 72 mm. This value is 99% of the sum for Japanese females. Judging from the above data, the vertebral columns of Minato gawa females may be somewhat shorter than those of the Recent Japanese females.
Data of the ninth thoracic and third lumbar vertebrae of MI indicate that his vertebral column might be equal in height to those of the Recent Japanese males (Tables 4. 3, 5). The heights of the second and fourth lumbar vertebrae from MIV. suggest that this individual had a slightly shorter vertebral column compared with Japanese females.
Thickness of the vertebral column
According to the measurement in Tables 4. 4-6, the column of MII shows roughly the same thickness as the mean for Japanese males, which is fairly thick in contrast to his slender limb bones. The columns of MII and MIII are slender in proportion to their limb bones. MIV is also slender in overall size but stout in appearance.
RIB
Materials
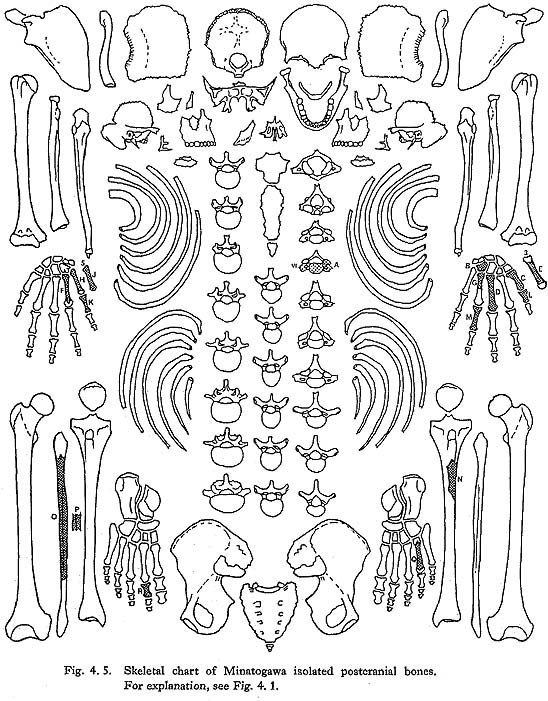
There remain more than one hundred small pieces of ribs. Unfortunately, however, only sixteen incomplete specimens have been identified (Figs. 4. 1-4, Plates 4. 5, 6). Namely, from MI the left sixth, the left eighth and ninth, the left eleventh. From MII, the left second and the right eleventh. From MIII the right first, the left fourth to sixth, the right seventh to ninth, the right eleventh and the left twelfth. From MIV, only the left second.
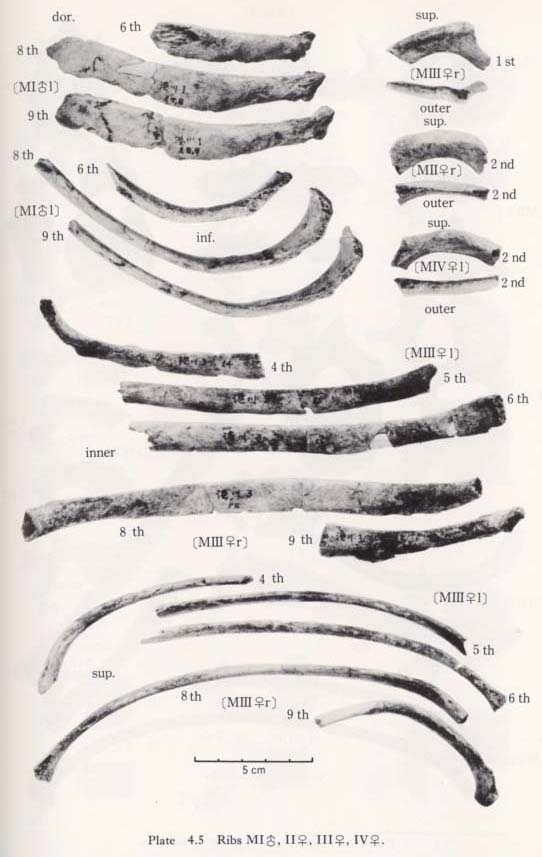
First and Second Ribs
There is only one first rib from MIII. Its head and the sternal half are missing. The tubercle is well developed. There. is a sharp ridge along the outer margin on the upper surface. The area medial to this ridge forms a shallow groove which may indicate the attachment of the scalenus medius. The width and thickness at the estimated midpoint are equivalent to those for Japanese females (Table 4. 7).
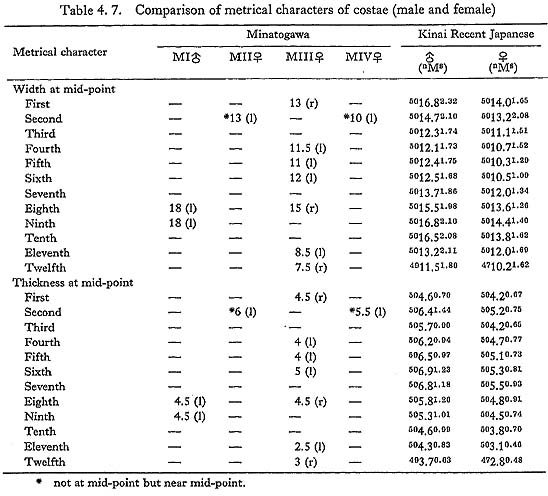
There are two second ribs from MII and MIV. The right bone of MII is a small fragment near the central but slightly vertebral part. On its upper surface, there is a sharp crest along the median line. A marked intercostal impression is seen on the inner half of the upper surface (Fig. 4. 7-A). By this crest and impression the cross-section in this region becomes flattened triangular in shape, resembling the feature of  V (McCown and Keith, 1939) and it is apart from that of the usual Recent Sapiens. On the distal end of the upper surface, a tuberosity which may be for the attachment of the scalenus anterior is obvious. The width and thickness near the midpoint are comparable to those at the mid-point for Japanese females.
V (McCown and Keith, 1939) and it is apart from that of the usual Recent Sapiens. On the distal end of the upper surface, a tuberosity which may be for the attachment of the scalenus anterior is obvious. The width and thickness near the midpoint are comparable to those at the mid-point for Japanese females.
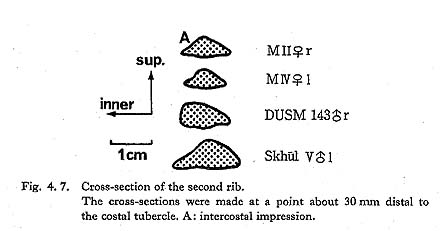
The remaining left bone from MIV occupies the vertebral part, but its head is missing. It also has a longitudinal ridge as in MII on the upper surface. The tuberosity for the scalenus posterior is well defined. Its width and thickness at the proximal part (slightly vertebral from the mid-point) are in the usual range for the Recent Japanese.
Middle Ribs from MI
Of these three bones, the larger two are similar in curvature, size and muscle marking. Accordingly, they must be adjacent bones. In addition, the head articular facet of the lower fits well with the corresponding facet of the vertebra, which is considered to be the ninth thoracic. Therefore, these two ribs may be the eighth and ninth. Another remaining rib seems to be the sixth thoracic. Their appearance is stout having strong muscle marking, particularly on the dorsal surface between the head and the angle. According to the measurement at mid-point, they are slightly wider and thinner than the corresponding ribs of Japanese males (Table 4. 7).
Middle Ribs from MIII
The right eighth rib is rather well preserved; only its head and neck are broken off. The estimated arch length of this bone is 25 cm, being equal to the mean for Japanese females. However, its chord length is estimated to be 20 cm, which is extraordinarily long. This very long chord might be due to the soil pressure. If not, this individual might have had a fairly narrow thorax. The height (width) and thickness of this bone at the mid-point are 15 mm and 4.5 mm. The height seems to be very large, while the thickness is normal (Table 4. 7). Other ribs show the same characters as that of the eighth.
Fragment of the Middle Rib from MIV
This is a small fragment, about 5 cm in length. Although it is not known to which rib it belongs, it is wide (17 mm) and rather thick (5.5 mm) as a female's would be.
SCAPULA
Materials
The following five scapulae were obtained, which belong to three individuals (Figs. 4. 1-4, Plates 4. 8, 9, 11).

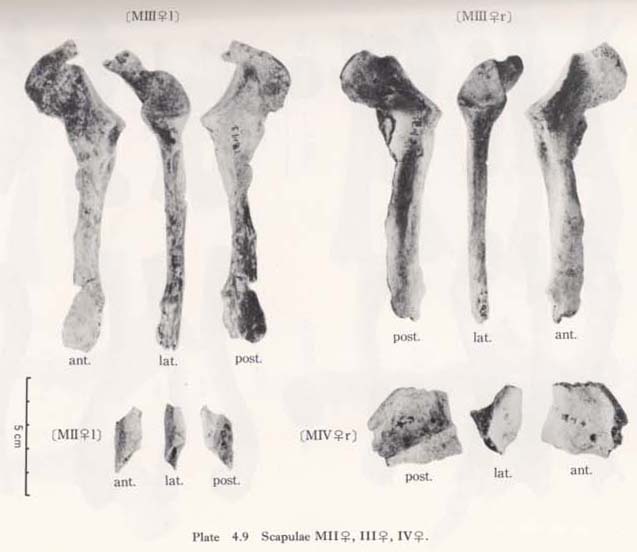

MIr: male, consisting of the glenoid, the axillary border and most of the spine and coracoid.
MI1: male, fragments of the axillary border and part of the spine.
MII1: small fragment of the axillary border.
MIIIr: female, consisting of the glenoid, the axillary border and part of the coracoid.
MIIIl: female, consisting of the glenoid, the axillary border, most of the coracoid, part of the spine and a small part near the vertebral border.
MIVr: female (?), fragment of the spine.
General Character
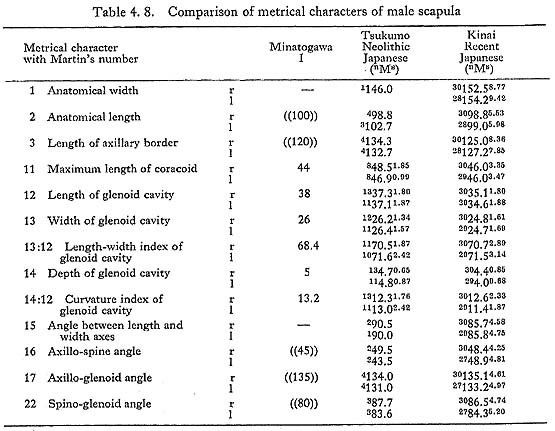
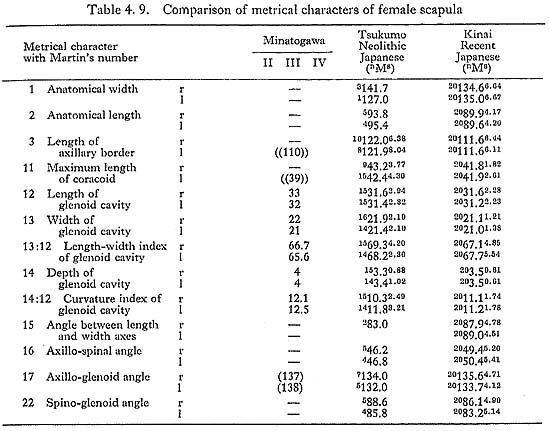
It is impossible to measure the whole size of the scapula because the specimens are fragmental. However, according to the remaining parts of the spine and axillary border, the anatomical length is estimated to be around 10 cm in MIr and the length of the axillary border is also estimated around 12 cm in MIr and 11 cm in MIIIl (Tables 4. 8, 9). The anatomical length of the MIr seems to match with the mean of Japanese males, while the length of the axillary border seems to be less.
Glenoid Cavity
The glenoids of Minatogawa scapulae are about the same in size as those of the Neolithic Japanese and rather larger than those of the Recent Japanese (Tables 4. 8, 9). The length-width index of all these specimens is slightly less than the means for the Recent and Neolithic Japanese. The length-depth index is 13 in MIr, equal to the means for the Japanese males. It is 12 and 13 in MIIIr1. They are slightly larger than those for the Japanese females.
As in usual cases of Japanese, the glenoid of Minatogawa scapulae face dorsally (Fig. 4. 8). The angle of the dorsal deflexion from the direct lateral position (McCown and Kieth, 1939) is estimated to be around 20° in MIr, 15° in MIIIr1. The long axis of the glenoid cavity is parallel to that of the axillary border. The anterior margin of the glenoid is thin but the posterior one is thick and round, as is also usual in the Japanese. The upward deflexion of the glenoid, i.e. the axillo-glenoid angle (135°-138°), is also within the range of Japanese. The supraglenoid tuber for the biceps brachii is well developed forming a round smooth surface in all the present specimens.

Spine
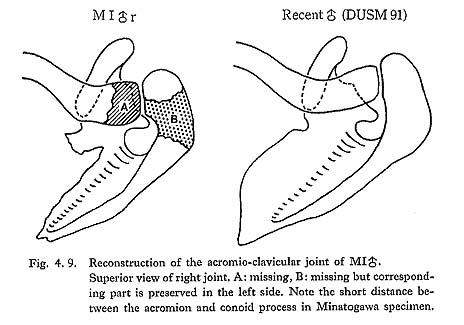
The spine of MI seems stout in proportion to its overall size. The thickness of the lateral margin at the mid-point is 7 mm in the right, 6.5 mm in the left. The spine decreases gradually in thickness as it comes medially. In most of the Recent Japanese, the lateral margin is thicker than that of MI, but the thickness decreases suddenly as it comes medially. The development of the acromion is not sufficiently observable because of partial breakage (Fig. 4. 9). Upward deflection of the spine seems to be normal. The distance between the glenoid margin of the spine and the rim of the glenoid is 17 mm in MIr.
Coracoid Process
The process is not thick but well projected. The distance between the top of the process to the nearest point of the glenoid margin is 25 mm in MIr. In MIl the root of the coracoid is rather thin and wide. Its lateral border is sharp and begins from the superior corner of the glenoid margin. In MIIIl, the root is thin and its lateral border is also rather sharp.
Axillary Border
The infraglenoid tubercle for the triceps brachii is well defined. From this tubercle, the lateral margin runs down along the middle line of the axillary border. After passing the mid-point it turns gradually posteriorly and fuses with the Crista dorso-axillaris (Vallois, 1932).
In other words, the Crista dorso-axillaris is situated medially, apart from the lateral margin (Fig. 4. 10). Accordingly, the Sulcus dorso-axillaris is present in the upper half of the axillary border. It is clear in MIr1 but rather faint in MIIIr1. The Crista ventro-axillaris in these specimens is of the usual sapiens type. The same condition can also be seen on the left small fragment of the axillary border of MIIl. The axillary border is rather thin, the thickness at the point 30 mm below the glenoid is 11 mm in MIr, 10 mm in MIIIl.

Fragment of the Left Spine of MIVl
lThis consists of a middle part of the spine, 37 mm in length and 33 mm in height including the base plate. The thickness of the glenoid border at the mid-point is 6 mm. The height of the spine from the base at a point 20 mm medial to the glenoid border is 26 mm, which is equal to the value for the corresponding height of MIl. Hence, there is a strong possibility that this fragment belongs to a male.
Discussion and Conclusion
The Minatogawa scapulae seem to have several characters comparable to those of the Neanderthals. However, the posterior deflexion and narrowness of the glenoid are also fairly common in the Recent Japanese. As for the development of the Crista dorso-axillaris combined with the Sulcus dorso-axillaris, Minatogawa scapulae belong to the Chancelade type, that is the intermediate between the Classic Neanderthal type and the Recent Sapiens type. It is well known that this type of axillary border is most common in the Upper Paleolithic Sapiens, and it is also seen in Recent Japanese (30% according to Takano, 1958).
CLAVICULA
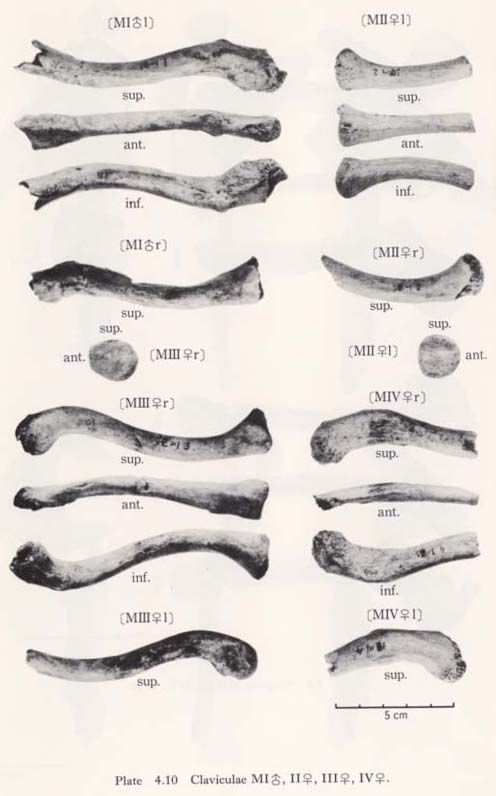
Materials
There remain eight bones, belonging to four individuals, as follows (Figs. 4. 1-4, Plates 4. 10, 11):
MIr: male, both ends are broken.
MIl: male, both ends are broken.
MIIr: female, medial half is preserved.
MIIl: female, medial half is preserved.
MIIIr: female, almost complete, a part of the acromial end is broken.
MIIIl: female, medial end is broken off.
MIVrl: female, both are lateral halves.
General Character
One of the most striking features characterizing the Minatogawa clavicle is its relative shortness. The maximum length in MIrl is estimated as 122 mm, which is extraordinarily smaller than the means for the Neolithic and Recent Japanese males (Fig. 4. 11, Table 4. 10). The maximum length of MIIIr shows the same characteristics as in MI (Table 4. 11).
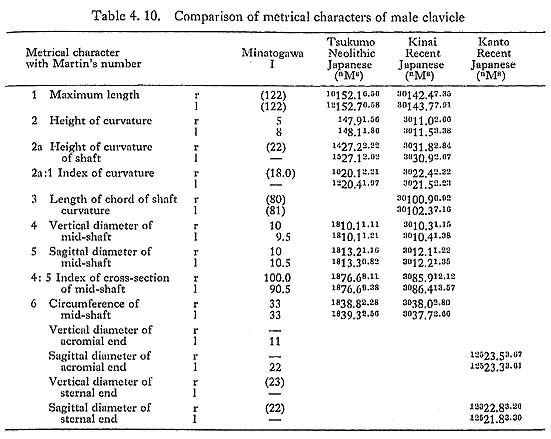
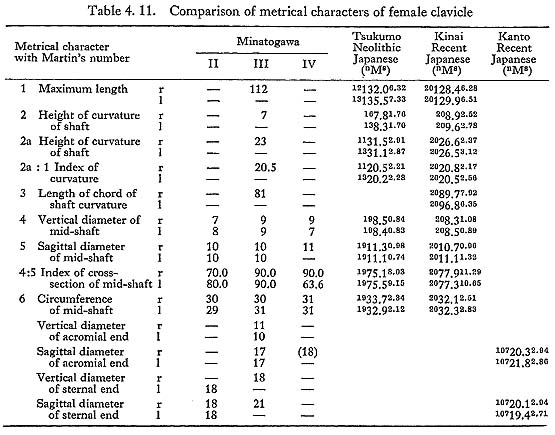

Their curvature indices are roughly equivalent to the Japanese. The circumference at mid-point is 33 mm in MIrl and varies from 29 mm to 31 mm in the female specimens. These values are slightly smaller than those of the comparative populations. The value of the length-thickness index is considerably larger in MIrl and MIIIr than the means in those populations.
Sternal End
The size of the sternal end in MIr and MIIIr is comparable to the mean for the Recent Japanese, while the value for MIIl is less than that of the Recent Japanese females. On the inferior surface, the impression for the costo-clavicular ligament is well developed forming a marked tuber in all the specimens.
Shaft
The shape of the cross-section at the mid-point is round in MIrl and MIIIrl, while it is flattened (platyleidic) in MIIrl and MIVrl. The shaft is not thick but is very short, and these specimens therefore seem stout. In MIrl, the impression for the pectoralis major and the groove for the subclavius are marked.
Acromial End
As commonly seen, the acromial end is flattened in all the Minatogawa specimens. In MI and MIII, it is very short and the conoid tubercle is well developed to form a prominent process. The trapezoid ridge is also marked. It is rather long and the tubercle and the ridge are not well developed in MIV.
The distance between the center of the conoid tubercle and the acromial facet is 30 mm in MIr, 29 mm in MIIIr. These distances approximately correspond to the distance between the conoid tubercle and the articular facet on the acromion. Accordingly, in MIr and MIIIr the acromion might be very long and curved anteromedially because the conoid tubercle in these cases is normally located (Fig. 4. 9). On the contrary, this distance is large in MIV (38 mm), the tubercle and the ridge are not well developed as in the case of Recent Japanese. The well-developed deltoid tubercle is seen in MIrl. In MIIIrl and MIVrl, an imprint or narrow fossa is seen instead of the tubercle.
Discussion and Conclusion
As mentioned above, the Minatogawa claviculae are relatively short. The claviculo humeral index is 42.5 in MI and 40.7 in MIII. According to the textbook of Martin (1928), these values are close to those of the Australians ( : 42.3) and Andamans (: 42.7, : 40.8) but vary greatly from those of the Japanese (: 49.6, : 47.9) and the Europeans (: 44.3, : 45.0). As far as MI is concerned, the clavicle is short in relation to the morphological length of the scapula. Therefore, it is likely that his shoulder was situated rather anteriorly; in other words, he had round shoulders.
There is a problem about the sex of MIVrl. The estimated total length and the anteroposterior diameter are larger than those of MI. Hence, it seems that these bones very probably belong to a male.