





コリドラス属 Corydoras
脊椎動物門硬骨魚綱カリクティス科
ブラジル
浦野貴士
1989〜90年採集
総合研究資料館、水産動物部門
一個体の標本も、一編の論文もない状態からこの研究は始まった。あったのは材料に対する強い好奇心だけである。
南米産の魚類は、熱帯魚としてかなりの量や種類数が日本に輸入されて来る。魚類「学」に何の興味もない方々にこれだけ知られ、魚類「学」のプロにこれほど知られていない魚も珍しい。思えば、これらの魚ほど危うい立場に立たされている生き物もいない。
南米のナマズを語るとき、日本の、長くて立派な髭をたくわえた、ぬるりとしたナマズを強く意識してほしい。知れば知るほど、あまりの違いに、それまでの固定観念があっけなく吹き飛ぶ興奮を味わうことが出来るかもしれない。その形、その仕草、その体に秘められた進化の謎を、1つ1つ明らかにしていけば、ひょっとするといつしか底無し沼から浮かび上がることが出来るかも知れない。
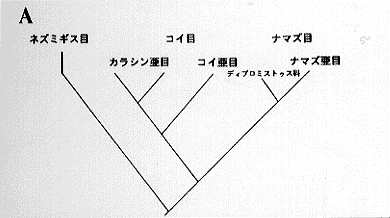
分布域は全ての大陸に及ぶ。同じウェーベル器官をもつコイ目が、南米やオーストラリアに分布せず、同様なカラシン目がユーラシア、オーストラリアに分布しないことを考えると、ナマズ目の広がりは全世界的なのである (Nelson, 1984)。
 | 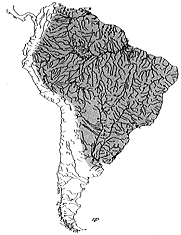 |
| 30-1 南米産ナマズ目魚類全科とそのアウトライン(−−、1994)。 | 30-2 カリクティス科の分布域。黒丸はコリドラス属の分布を表す。 |
ただ種類数が多いだけではない。形態的特徴にも様々な変化があり、多様性は極に達している。ここにはもはや、日本の我々に馴染みのあるナマズは存在しない。
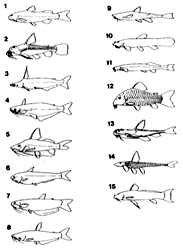
(1) ディプロミストゥス科 Diplomystidae 2属3種。チリとアルゼンチンに限産する。最大28センチ。ナマズ目中唯一、発達した主上顎骨に多数の歯と一対の髭を持つ。6枚の下尾骨を持つ。2〜4番までの脊椎骨が融合し、単純なウェーベル器官を持つ。現在知られているナマズ目魚類のなかで、最も祖先的形質を保持している一群であり、ナマズ目の進化を考える上で重要な位置を占めている (Arratia, 1990)。
(2) ドラス科 Dordidae 約35属90種。主にブラジル、ペルー、ギアナ。最大約80センチ。体側に大形の骨板と3対の髭を持つ。背鰭、胸鰭の棘条はよく発達する。この胸鰭棘条と肩帯の骨をこすり合わせて発音させ、鰾で増幅し大きな威嚇音を出す。脊椎は7番まで融合する。
(3) アゲネイオスス科 Ageneiosidae 2属約12種。パナマ。南米熱帯域からアルゼンチン。最大100センチ。偏平した頭部とごく短い一対の髭を持つ。無鱗。脂鰭は小さい。
(4) アウケニプテルス科 Auchenipteridae 21属約60種。パナマ。南米熱帯域からアルゼンチン。2〜3対の髭を持つ。無鱗。脂鰭はないが、あっても小さい。ある種では、産卵時、雄の背鰭棘や髭が変形し、雌を締め付ける道具に変化する (Curran, 1989)。
(5) ピメロドゥス科 Pimelodidae 約56属300種以上。南米全域、南部メキシコ。ナマズ目中最多の種数を持つ。最大300センチに成長するブラキプラティストーマ・フィラメントースム Brachyplatystoma filamenosumは、淡水魚類中最大魚種の1つ。南米で最も繁栄しているグループであり、食料として重要な位置を占める。
3対の髭と脂鰭を持つ。無鱗。背鰭、胸鰭の棘条は多くの種が持つが、ないものもある。大型の種は高い遊泳能力を持ち、河川、湖沼の低層域だけでなく、広い遊泳域を確保し、汽水域にも生息範囲を広げている (Schaefer, 1990)。
(6) セトプシス科 Cetopsidae 4属約12種。南米全域。3対の髭を持つ。無鱗。脂鰭もない。鰾は退縮的で骨のカプセルに包まれる。貪欲な肉食性を示す、南米で最も恐れられている魚種を含む。軟らかい唇で体に吸いつき、発達した歯で噛みきる。目や髭は退化的。高い遊泳能力を持つ。
(7) ヘロゲネス科 Helogeneidae 1属4種。南米熱帯域。最大10センチ。3対の髭を持つ。脂鰭はあっても小さい。背鰭は後方に位置し、胸鰭とともに棘条はない。無鱗 (Vari and Ortega, 1986)。
(8) ヒポフタルムス科 Hypophthalmidae 1属3種。南米熱帯域。最大60センチ。3対の髭を持つ。無鱗。脂鰭は小さい。偏平な大きな頭部の下方に目が位置する。鰓孔は大きく、歯を欠く。遊泳しながら餌を濾し採るのではないかと考えられる (Howes, 1983)。
(9) アスプレド科 Aspredinidae 10属32種。南米熱帯域、一部の種は汽水域に生息する。最大42センチ、多くは15センチ未満。1〜2対の髭を持つ。体側に突起状の骨板が並ぶ。脂鰭はない。鰓孔は小さく水を噴出して前進したり、腹鰭を左右に動かし、歩くように移動する事ができる。多くの種で、背鰭の固定機能がない。胸鰭棘条は発達する (Ferraris, 1991)。
(10) ネマトゲニス科 Nematogenyidae 1属1種、チリに限産する。3対の髭を持つ。無鱗。脂鰭はない。トリコミクテルス科の亜科とされていたが、近年独立した。鰓蓋に棘がない。
(11) トリコミクテルス科 Torichomycteridae 約36属155種。南米全域、中南米。1〜2対の髭を持つ。無鱗。ドジョウ様の体形だが頭部は偏平。脂鰭はないものが多い。鰓蓋に棘を持つ。腹鰭を欠く種がある。セトプシス科の一種と同様、南米で最も恐れられている魚種を含む。他の魚類や動物の体に食い込んだり、鰓に食いつき吸血する。また、魚の分泌する粘液や鱗を食う種もある。体色はほとんど透明から、複雑な文様を持つものまで様々。渓流の砂利底から大河の泥底に生息する種までいる (Pinna, 1989)。
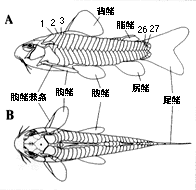
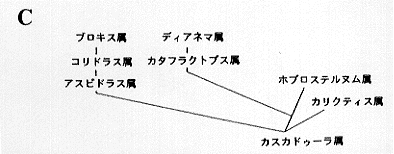
(12) カリクティス科 Callichthyidae 7属約140種。南米全域。最大12センチ。2亜科7属に分類される。カリクティス Callichthys 1種、カタフラクトプス Cataphractops 1種、ディアネマ Dianema 2種、ホプロステルヌム Hoplosternum 3種、ブロキス Brochis 3種、アスピドラス Aspidoras 14種、コリドラス Corydoras 124種が認められている(挿図3)。化石種としては、コリドラス1種が知られている。3〜4対の髭と体側に2列の大きな骨板を持つ。体は側偏、頭部はカリクティス亜科 Callichthinae の魚種では縦偏し、コリドラス亜科 Corydoradinae の魚種では側偏する。鰾は骨に囲まれ上後頭骨後部に付着する。口は小さく下方に開き、突出させる事が出来る。背鰭、胸鰭棘条は鋭く発達し、脂鰭は骨板により支持される。腸呼吸を行う。ナマズ目中最小の種コリドラス・ハスタートゥス C. hastatus、コリドラス・ピグマエウス C. pygmaeus(挿図4)を含む。コリドラス属は現生魚類中最大の属である。現在も新種の発見が相次いでおり、一説には200種を超えるのではないかといわれている(Nijssen & Isbr ker, 1970; 1975; 1979; 1980、挿図5〜7)。
ker, 1970; 1975; 1979; 1980、挿図5〜7)。
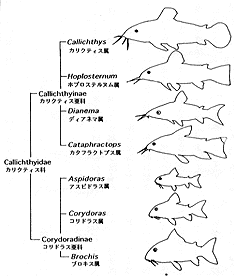 |  |
| 30-3 カリクティス科全属とそのアウトライン | 30-4 コリドラス属中最小種のピグマエウス(Corydoras Pygmaeus)と最大種のバルバートゥス(C.Barbatus)。 |
 | 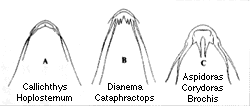 |
| 30-5 カリクティス科アスピドラス属の外部形態。A)側面図、数字は骨板数を示す。B)背面図。 | 30-6 カリクティス科7属の髭の形態。 |
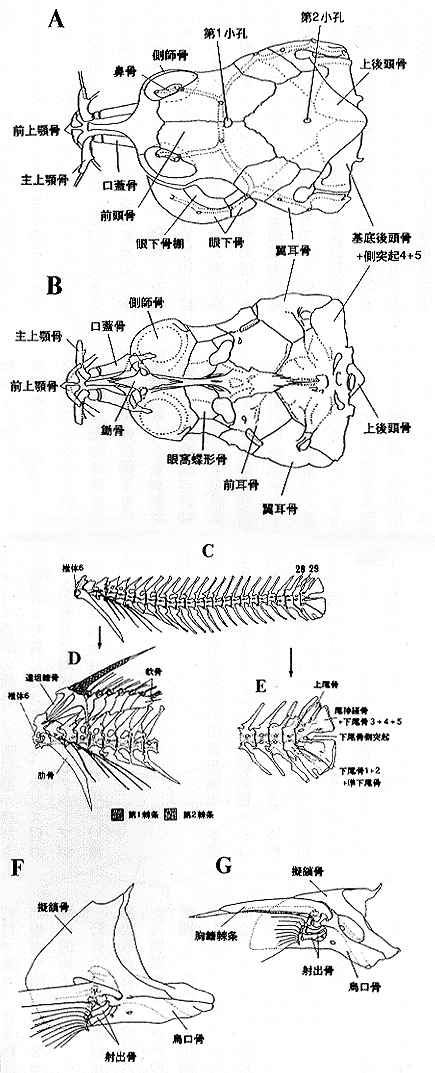 | 30-7 アスピドラス属の骨格。A)頭部背面図、B)頭部腹面図、C)脊椎骨、D)前部脊椎骨と胆鰭骨、E)尾鰭骨格、F)右}、G)肩帯腹面図 |
(13) スコロプラックス科 Scoloplacidae 1属4種。ペルー、ボリビア、ブラジルの一地域に限産。最大2センチ。4対の髭を持つ。体側や頭部に皮歯のある骨板を生じる。脂鰭、鋤骨、側後頭骨を欠く。背鰭、胸鰭棘条は発達する (Schaefer, 1990)。
(14) ロリカリア科 Loricariidae 約80属550種。南米全域。最大90センチ。骨で支持される髭を欠く。体は縦偏しほとんど骨板で被われる。脂鰭を欠くものの多くは、長く伸びた尾部を持つ。口は下部に開口し多くは肉質突起を伴った吸盤状をなす。背鰭、胸鰭の棘条は発達する。ロリカリア科魚種の目には光に感じ光量を調節する虹彩がある。
本科魚種は、南米で最も繁栄しているグループである。南米全域のあらゆる水域に出現しその数も多い。河口付近の淡水域から、標高3000メートル以上の高地の渓流域にまで分布を広げている。生態的にも重要な位置を占めると考えられる。また、大型のものは、食用としても重要である。
(15) アストロブレプス科 Astroblepidae 2属約40種。アンデス地域とパナマ。最大30センチ。2対の髭を持つ。無鱗かほとんど無鱗。口は円盤状で吸盤となる。脂鰭はあるものと欠くものがある。背鰭棘条は固定できない。いくつかの種類は山間部の激流の中でも生息しており、滝の裏側を登ることが知られている。
ker, 1970; 1975; 1979; 1980; 1983)。本学では、1985年頃より研究がスタートした。1988年の第1回ブラジル調査から毎年の調査により標本数も増加しており、ナマズ目では10科、カリクティス科魚類では6属約80種余りを確保している。また、生体を飼育しての初期発生の研究も並行して行っており、本科魚種の新たな側面が明らかになりつつある。
属ごとの分布は今だに明確ではない。コリドラス属が、最も広範囲に分布を広げていることは確かだが、調査が進めば、他属でも分布域が拡大して行く可能性は十分にある。近縁なアスピドラス属は、ブラジル高原を中心に分布していると考えられていたが、調査が進んだ事によりアマゾン本流域からも確認されている。調査が行き届かない原因としては、分布域が広範囲であること、未開発地域が多く残されていることが挙げられる。現在の資料ではカリクティス科魚類の分布域を正確に認識することは難しい(挿図2)。
前頭骨に一小孔があるが、アスピドラス属では上後頭骨にも一小孔を持つ、成長とともに、これは小形になり、個体によってはふさがれるものも観察された(挿図7A)。
鰾周辺の骨は著しく特化している。4と5番目の脊椎骨が融合し、生じていた側突起が骨嚢となり、後方から浮袋を覆い隠している。鰾そのものが腹腔内から押し出され、後頭骨後方に位置したため、側突起から生じた骨嚢と上後頭骨は間接しあい、頭骨の一部と化している。1〜3番までの脊椎骨は押しつぶされ基底後頭骨と融合している。ここに発生しているウェーベル器官は単純化し三脚骨とせん状骨は融合して鰾に接している(挿図7A、B、8)
6番脊椎骨に関節する肋骨は強大であり、先端から出た腱が体側の骨板に付着し体を固定している。後方の肋骨は極端に細くなる。脊椎骨から出る神経棘は幅広く変形し、背鰭担鰭骨が突き刺さっている。後方の神経棘にも同様の構造が見られる(挿図7C、D)。
背鰭棘条は2本あるが、第一棘条は小形のくさび状になり、第一棘条を固定するために働く。2つは腱で接続しており、第一棘条が担鰭骨に引き込まれるのに伴い、第二棘条が立ち上がる。第一棘条は湾曲しており、深く担鰭骨に入り込むと完全にロックされる(挿図7D、9)。
尾骨格を形成する下尾骨は上下2つに分かれ、上部では尾神経骨と3〜5番までの下尾骨が融合し、下部では準下尾骨と1、2の下尾骨が融合している(註20)。(挿図7C、E)咽頭歯は鋭い犬歯状で、よく発達する。
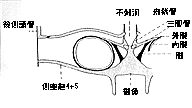 |  |
| 30-8 カリクティス科魚類のウェーベル器官模式図(Alexander、1964)。 | 30-9 カリクティス科魚類の背鰭ロック機構模式図(Alexander、1965)。 |
胸鰭を支える肩帯は単純化し、擬鎖骨と烏口骨が大きく発達する。胸鰭棘条の基部は鍵型に湾曲し、烏口骨の突起を引っかけるように回転する事が出来る。擬鎖骨は棘条が外れるのを防ぎ、かつ固定するのを助ける。ほとんどの種で棘条の内側に揃った鋸歯があり、カリクティス亜科の場合、この形態が種群を決める重要な形質になっている。ナマズ目の中にはこの部分に雌雄差が現れるグループがあるが、本亜科ではその様なことはない(挿図7F、G)。
南米の河川や湖沼は、雨期と乾期で大きく水位が変動する。そのため、水質や水温、水深、流量に多大な影響を与える。特にカリクティス科魚類の生息域は、その様な影響を比較的受けやすく、彼らは多様な環境に適応出来る能力を身につけている。腸呼吸の能力を獲得したことは、彼らの適応力を飛躍的に高めたと言える (Low-McConnell, 1987)。
多くのナマズ目魚類が夜行性なのに対し、本科魚類は主に昼行性である。体側を被う頑丈な骨板と棘条を備えたことで、補食者を退けることに成功した。なお、仔魚は完全な夜行性であり、光を極端に嫌うことが、水槽での観察で明らかになった。この性質は成長に伴い消失する。
野外での繁殖観察は報告されていないが、採集された個体サイズおよび飼育による成長から判断すると水深の浅くなる乾期はじめに産卵し、乾期から雨期にかけて仔稚魚が成長するようである。しかしこれを確信するためには、より多くの観察事例が必要である。
本科魚類の受精生態については諸説あったが、近年雌が精子を体内通過させ自らの卵を受精させることが明らかとなった。雌は雄の排出口に口を近づけ精子を吸い込み、その直後腹鰭を閉じて袋状にすると、そこへ卵を産み落とす。体内を通過した精子は、排出口から腹鰭に生み出されている卵に受精するのである。その後雌は、卵を基質に付着させるため泳ぎだす。アスピドラスなど、かなり流れの早い河川に生息する種でも、こうした受精方法であれば、確実に子孫を残すことが可能である。
孵化仔魚はおたまじゃくし様で、2対の髭、胸鰭と尾部を包む大きな膜鰭がある(挿図10)。目は不完全。鰭は、尾鰭、背鰭、尻鰭、腹鰭の順に鰭条が完成される。脂鰭は、膜鰭の取り残されたものであり、膜鰭中に生じた骨板により支持される。体側の骨板は鰭の完成後に遅れて生じる。始め頭部後方と尾柄部より出現し、体側中央に縦列する。その後背部、腹部方向に成長し、完成する。体表の色素胞は増加集合を繰り返しながら、種独特の模様を作り出す。目から吻端にかけての黒帯は、コリドラス属、アスピドラス属の特徴である。
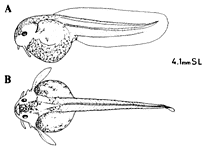 | 30-10 アスピドラスtスコグッタートゥス(AspidorasFuscoguttatus)の孵化仔魚。 |
この圧倒的種数のコリドラス属を、ナイスンとイスブリュッカーは体形、斑紋パターン、胸鰭棘条後縁の鋸歯形態などにより5つの種群に分けた(挿図11)。しかしながら、明らかに特徴のあるエレガンス・グループとアークトゥス・グループを除くと、妥当性は薄いと言える。特にプンクタートゥス・グループは、点模様の斑紋パターンを重要視したためか、ごった煮状態に近く問題が多い。
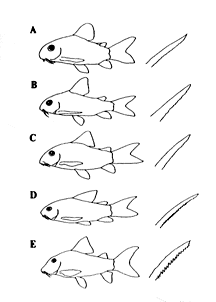 | 30-11 ナイスン、イスブリュッカーによるコリドラス属内の7種群のアウトライン。右は胸鰭棘条。A)プンクタートゥスOループ、B)アエネウスOループ、C)バルバートゥスOループ、D)エレガンスOループ、E)アークトゥスOループ。 |
1990年から1994年までに行ったアマゾン川の大支流、アラグアイア水系の現地調査で、極めて似かよった点模様を持つ、しかし明らかに別種と考えられる未記載種4種の分布パターンを明らかにした。4種の主な分類基準は次の通りである(口絵参照)。
(1) 体形、(2) 点模様の大きさ、(3) 点模様の入る位置、(4) 尾柄部骨板の数および配置、(5) 腹面に生じる骨板と棘の、存在および配置、(6) 間鰓蓋骨の体表に現れる割合(挿図12〜14)。
この地域を流れる同水系には、既知種2種アラグアイアエンシス C. araguaiaensis とマクリフェル C. maculifer が知られており、これらも同様な点模様を持っている。文献ナイスンらの分類基準では、マクリフェルはアークトゥス・グループに入るがアラグアイアエンシスと未記載種4種は、全てプンクタートゥス・グループに分類される(口絵参照)。
これら6種の分布を調査すると、実に興味深いことが分かる。アラグアイアエンシスとsp. 3はアラグアイア川支流のゴヤス州側と、同川の支流の1つであるモルテス川右岸に流れ込む小河川から広く採集された。sp.1、sp.2は、両河川の間を流れる支流のみから採集され、sp.4は、アラグアイア川上流域の1ケ所から報告された。マクリフェルは、モルテス川右岸に注ぐ数本の小河川より採集されている(挿図15)。
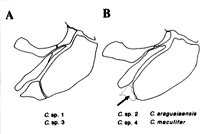 | 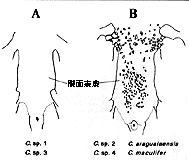 |
| 30-12 コリドラス属鰓蓋部の2パターン。A)間鰓蓋骨が全て露出する。B)間鰓蓋骨(点線部)は表皮に覆われる。 | 30-13 コリドラス属腹部の2パターン。A)骨板は生じない。B)骨板が生じる。 |
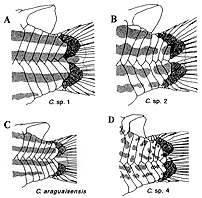 | 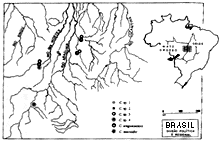 |
| 30-14 コリドラス属尾柄部の模様と後端骨板配列 | 30-15 コリドラス属6種の分布 |
この地域を流れるアラグアイア川とモルテス川は、川幅100メートル以上、乾期には10メートル近く水位が変動する年もあるとは言うが、それでも流量、流速、コリドラスの遊泳能力や性質を考えると、とても生息域を広げることは出来ないと思われる。アラグアイアエンシスの広い分布域は、これらの河川が今の状態になる以前に形成されたのであろう。それではsp.1、sp.2はどうなのだろうか。調査地域が限定されているため、分布の広がりを把握しかねるが、分布が限られる種である可能性は高い。マクリフェルも分布の範囲はかなり限られ、アラグアイア本流側には生息しない。このように種によってその分布状態はかなり異なることが明らかになった。
さらに同一地点での生息状態を見ると、非常に似通った斑紋パターンを持ちながら、吻端の長い種と短い種がともに生息している。アラグアイアエンシスとsp.3、sp.1とsp.2がこの組合せに当たる。マクリフェルの生息場所では、アラグアイアエンシスとsp.3も、ともに生息していた。同一地点に3種類のコリドラスが、それもかなりの個体数生息していたのである。
このような例は、コリドス属では時折報告されている。これを雌雄差ではないのかとの疑問もあるようだが、sp.3の吻端を短くしたタイプであるアラグアイアエンシスで見る限り、雌雄が存在し繁殖した事から、sp.3の中にも雌雄は存在し、独立した種であると考えている。ただ吻端の長いタイプは個体数がかなり少なく、単独で採集されたことが無いことが気にかかる。マクリフェルなど、アークトゥス・グループの種にも同様の事が言える。個体数は吻端の短い種よりかなり少ない。これら3タイプの種は同一地域ではともに混群を形成し、ほぼ同一の体長を持つ個体どうしが、数尾から十数尾の群れを作り浅い水底で盛んに採食活動を行っていた。水槽内での観察では、別種どうしが雑種を作ってしまうことも度々報告されており、これらの非常に似通った斑紋パターンを持つ種どうしが同一地点に生息している場合、どの様に繁殖しているのか知りたいところである。このような例は数千キロ離れたアマゾン水系各地で発見されており、圧倒的多数を誇るコリドラス属の種分化や分布、分類を考察する上で大変興味のある問題となっている (Burgess, 1982; Fraser-Branner, 1947; Nijssen & Isbrker, 1983)。
 | 30-16a ノヴァセック・マーシャル、1976の提示した骨鰾類の系統関係 |
 | 30-16b ハウス、1983の提示したロリカリア科その他の系統関係 |
 | 30-16c ゴスライン、1940の提示したカリクティス科の系統関係 |
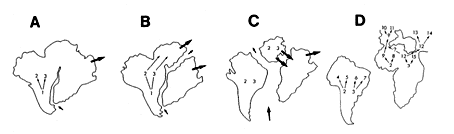
30-17 南米大陸とアフリカ大陸の分裂と移動(ノバセックとマーシャル、1976)。
A)白亜紀初期、B)白亜紀中期、C)白亜紀中後期、D)白亜紀後期〜暁新世。
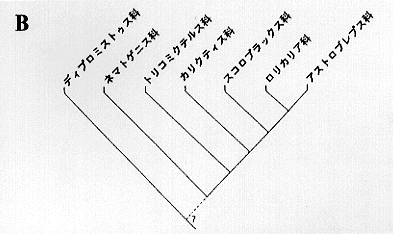
ナマズ目を網羅する系統関係を打ち出したのはシャルドンである。彼は脊椎骨や肩帯、浮袋の形態により推定を行ったが、形質の種類数や組合せに問題があり、多くの支持を得られず現在に至っている。しかしカリクティス科に関して言えば、ロリカリア科、アストロブレピス科、トリコミクテルス科等と近縁としており、他の研究者とも意見が一致する。さらにハウスは、ネマトゲニス科、スコロプラックス科を独立した科と認め、これらを含めた類縁関係について論議し、ロリカリア科と共通の形質を多く持ちながらも、スコロプラックス科をより近縁なグループとしている(挿図16B)。
カリクティス科内の系統関係については、ゴスラインの研究があるが、ホプロステルムヌ属の幼魚をカスカドゥーラ属 Casucadura としてしまうなど、分類学的に問題を含んでいる。(挿図16C)それ以後、新しい系統関係の推定は成されていない。
近年、生物の分布を大陸移動と関連づけて説明することはごく一般的になった。ナマズ目の分布を考える上で、ゴンドワナ大陸の存在と大陸移動の事実は、大変便利な材料である。南米、アフリカと続き、東南アジアがゴンドワナの一部であるとすれば、この地域において華々しく分化、放散をとげたナマズ類の進化も合理的に説明がつくかも知れない。ディプロミストゥスの祖先がナマズ目魚類の共通の祖先であることはだれしも認めるところだが、それに続く種がハマギギ科のような塩分耐性を獲得していたとするなら、現在の分布を考える上でまた新しい展開をみせるだろう。いずれにせよ、地史の解明とともに、信用に足る系統類縁関係の構築が不可欠であり、その基礎に分類学が置かれていることは言うまでもない。
(浦野貴士)
 flexions sur la dispersion des Ostariophysi
flexions sur la dispersion des Ostariophysi  la lumire de recherches morphologiques nuvelles. Ann. Soc. R. Zool. Belg. 97 (3): 175-186.e de l'appareil de Weber et des structures connexes chez les Siluriformes. Mus. R. Afr. Cent. Ann. (Ser. 8, Zool.) 169: 1-277.
la lumire de recherches morphologiques nuvelles. Ann. Soc. R. Zool. Belg. 97 (3): 175-186.e de l'appareil de Weber et des structures connexes chez les Siluriformes. Mus. R. Afr. Cent. Ann. (Ser. 8, Zool.) 169: 1-277. ker. 1970. The South American catfish genus Brochis Cope, 1872 (Pisces, Siluriformes, Callichthyidae). Beaufortia, 18 (236): 151-168.ker. 1975. Cataphractus punctatus Bloch 1794 (Pisces, Siluriformes, Callidhthyidae): request for invalidation of neotype and validation of a rediscovered syntype as lectotyp. Z.N. (S.) 1950. Bull. zool. Nomencl., 32 (1): 63-64, pl.l.ker. 1979. Chronological enumeration of nominal species and subspecies of Corydoras (Pisces, Siluriformes, Callichthydae). Bull. Zool. Mus. Univ. Amst. 6: 126-135.ker. 1980. Areview of the Genus Corydoras Lac
ker. 1970. The South American catfish genus Brochis Cope, 1872 (Pisces, Siluriformes, Callichthyidae). Beaufortia, 18 (236): 151-168.ker. 1975. Cataphractus punctatus Bloch 1794 (Pisces, Siluriformes, Callidhthyidae): request for invalidation of neotype and validation of a rediscovered syntype as lectotyp. Z.N. (S.) 1950. Bull. zool. Nomencl., 32 (1): 63-64, pl.l.ker. 1979. Chronological enumeration of nominal species and subspecies of Corydoras (Pisces, Siluriformes, Callichthydae). Bull. Zool. Mus. Univ. Amst. 6: 126-135.ker. 1980. Areview of the Genus Corydoras Lac pde, 1803 (Pisces, Siluriformes, Callichthyidae). Bijdragen tot de Dierkunde, 50(1): 190-220.ker. 1983. Septespces nouvelles de Poissons-Chats cuirass du genre Corydoras Lacepde, 1803, de Guyane fran
pde, 1803 (Pisces, Siluriformes, Callichthyidae). Bijdragen tot de Dierkunde, 50(1): 190-220.ker. 1983. Septespces nouvelles de Poissons-Chats cuirass du genre Corydoras Lacepde, 1803, de Guyane fran aise, de Bolivie, d'Argentine, du Surinamet du Bresil (Pisces, Siluriformes, Callichthyidae). Revue fr. Aquariol., 10 (3): 73-82.
aise, de Bolivie, d'Argentine, du Surinamet du Bresil (Pisces, Siluriformes, Callichthyidae). Revue fr. Aquariol., 10 (3): 73-82.