




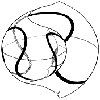

アクタウオ Lactarius lactarius (Block and Schneider)
脊椎動物門硬骨魚綱真骨類アクタウオ科
アンダマン海、横断面、寒天法(註1)により作成
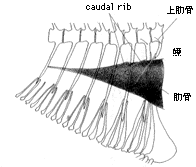
体の右側を走る後方延長部をもつ鰾ZUMT(註2) 59696
総合研究資料館、動物部門
「脊椎動物は淡水域に起源があり、初期の魚類の進化は淡水域で起こった。したがって、硬骨魚類も淡水域に出現し、その後海洋に進出した」と長い間考えられてきた (Romer, 1968)。しかしながら、現在では、多くの化石の証拠から、「脊索動物から脊椎動物への進化は、古生代のはじめ頃(カンブリア紀後期、約5億2000万〜5億500万年前)に、浅海域で起こり、したがって初期の魚類の進化の舞台は海であった」と考えられている (Carroll, 1987; Long, 1993)。最初に出現した脊椎動物は顎を持たない無顎魚類であるが、デボン紀(約4億800万〜3億6000万年前)になるまでには、全ての主要な有顎魚類(板皮類、棘魚類、軟骨魚類、硬骨魚類)が次々と海に出現した (Carroll, 1987; Long, 1993; Denison, 1978; Zangerl, 1981)。それでは、古生代に出現した硬骨魚類は、その後どのような進化の道を歩んだのであろうか。
ここでは、硬骨魚類に特有の器官である鰾の形態と機能からこの問題を考えてみたい。
鰾の最も一般的な機能は魚体の浮力調節であるが、この他呼吸、聴覚補助、水圧受容器や発音器官としても作用することが知られている。
ところで、4足動物の呼吸器官である肺と魚類の鰾は、消化管の一部が膨れて生じた相同器官と考えられている(挿図1、2)。魚類から4足動物へという脊椎動物の進化からみると、鰾が肺に変化したと推測されがちであるが、実際にはこの逆のようである。そして、この肺から鰾への変化とその後の形態の変化には、生息場所の移動による環境の変化が大いに影響していたらしい。
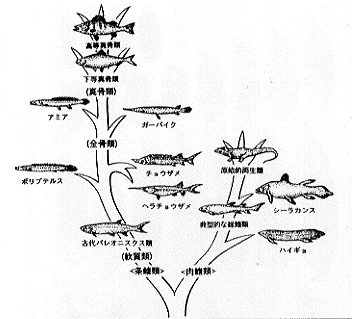 | 29-1 硬骨魚類の系統樹(Romer, 1959を改変) |
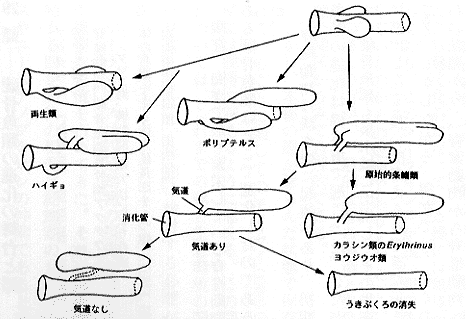 | 29-2 鰾の系統発生(Jollie, 1973を改変) |
化石の記録によれば、古生代のシルル紀(約4億3800万〜4億800万年前)の海に出現した初期の硬骨魚類はすぐに汽水や淡水域に進出し (Carroll, 1987)、デボン紀(約4億800万〜3億6000万年前)の淡水域で繁栄した (Romer, 1959)。デボン紀の原始的な条鰭類であるパレオニスクス類などは、既に肺のような構造を持っていたと考えられている (Colbert and Morales, 1990)。もちろん、空気を満たした肺は、鰾と同じように魚体の比重を減少させる (Alexander, 1975)。
この肺の出現には、デボン紀の気候条件が深く関係していたらしい。魚類の時代とも呼ばれるデボン紀は気候の変動が激しく、現在の熱帯域の一部のように、雨期と乾期の交代があった。乾期が始まると川や池の水は停滞し、鰓呼吸に必要な水中の酸素が不足する。この時に、毛細血管が密に分布している上皮を持つ肺のようなものが発達していれば、大気中の酸素を利用することができたであろう。肺は硬骨魚類が当時の乾期を生き抜くための適応の産物であったと考えられる (Romer, 1959; Romer and Parsons, 1977)。
現生種も含むハイギョ類は、デボン紀のはじめ頃浅海域に出現し、デボン紀中期までには淡水域に進入し始めた (Long, 1993; Campbell and Barwick, 1986)。現在もオーストラリア、アフリカおよび南アメリカの熱帯地方の淡水域に生息しているハイギョ類では、肺がよく発達している。さらに、デボン紀以降に出現した原始的な条鰭類で、現在アフリカの熱帯の河川に分布する軟質類のポリプテルス類や北・中部アメリカの淡水系に生息する全骨類のアミアやガーの仲間も肺のような構造を持ち、ある程度空気呼吸をすることが知られている。
このような肺を持つ硬骨魚類の中で、柄のある鰭を持つ肉鰭類から両生類が進化した(註3)。
デボン紀の初期には、魚類化石の記録は主として淡水産のものであったが、時代が進むにつれて、海産種の割合が増加している。これは魚類の淡水域から海洋へと移住する傾向が強まったことを示している (Romer, 1959)。
ところで、両生類、ハイギョ類、軟質類(ポリプテルス類やチョウザメ類など)では、肺・鰾と消化管を連結する気道(気管)は消化管の腹面に開くが、全骨類や真骨類では消化管の背面に開く。いずれの場合でも、魚類では肺・鰾が背方にあるのは水中における体の安定性のためであることは明らかである。この気道の位置については、真骨類のカラシン類の Erythrinus やヨウジウオ類などに中間的なものがみられる(Jollie, 1973、挿図2)。
中生代(約2億4800万〜6500万年前)になると、条鰭類の多くのものは海への移住を完了した (Romer, 1959)。今日最も繁栄している真骨類は、軟質類より進化した全骨類を経て、中生代のはじめ頃(三畳紀)に出現した (Carroll, 1987)。デボン紀の不安定な環境と異なり、海洋は安定しており、空気呼吸(補助的あるいは緊急用の呼吸器官としての機能)の必要がなくなり、肺は完全に浮力調節器官としての鰾に変化した。
その後、真骨類は沿岸から沖合へ、表層から中・深層へ、浅海底から深海底へとあらゆる海洋環境に進出した。様々な環境への適応に伴って、鰾の形態は著しく多様化した。これらの多様性は、それぞれの魚類の遊泳能力や生息環境への適応の様式などの違いによって生じたものである。たとえば、表層を遊泳するニシン類、浅海に生息するスズキ類、海底付近を遊泳するタラ類などでは、鰾はふつうよく発達している。しかし、ハゼ類・カサゴ類の多くの種、カレイ類の成魚などの底生魚やコバンザメ類やマンボウ類などの活発に遊泳することのない魚は、鰾を欠くようになった。1000メートル以深に生息する深海魚では、鰾は水圧の変化の影響が大きく、かえって生活の邪魔になるので、一般に退化的か失われてしまっている。
真骨類の中で原始的なニシン類、コイ類、サケ類、ウナギ類などでは、成魚になっても気道によって、鰾と消化管は連結しているが、高等なスズキ類やタラ類などでは、発生の初期に気道が消失して、鰾の一部にある特別な組織(赤腺と卵円体)を通して気体の増減を行う。現在、淡水域において最も繁栄している真骨類であるメダカ類、カラシン・コイ・ナマズ類は、一般によく発達した鰾をもっている。これらの魚類は再び淡水域に戻ったものである。ナマズ類のゴンズイの仲間などは現在浅海に進出している。また、南アメリカのカラシン類の Erythrinus の仲間など一部の淡水性の真骨類では、鰾が空気呼吸の機能を持っている。
以上のように、硬骨魚類は海に起源し、その後、海から淡水域、淡水域から海、そして一部は再び淡水域へと進化の舞台を変えていったのである。このような生息場所の移動による環境の変化に伴って、肺と鰾はそれぞれの環境に適応するために、その形態と機能を様々に変化させていったのである。
ところで、真骨類のいくつかのグループでは、鰾が腹腔の後端より後方へ延長することが知られている。
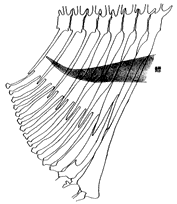
著者は、総合研究資料館において共同研究者と、スズキ亜目魚類(註4)の系統分類学的研究の一環として、本亜目魚類の鰾の後部の形態とそれに関連する構造を観察してきた(註5)。その結果、スズキ亜目の多くのグループでは鰾は腹腔に収納されているが、いくつかのグループの鰾は常に後方への延長部をもつこと(口絵、挿図3〜7)、中には鰾を欠くグループもあることが判明した。次に、本亜目内の類縁関係の推定において、この後方への延長という形質がはたして有効であるかどうかについて検討した。既に述べたように、鰾の形態は遊泳能力や生息環境と強い相関が想定されるため、適応的な形質とみなされ、したがって類縁関係の推定には不適と考えられるかもしれない。しかしながら、同じような延長の様式(その度合、左右相称性など)や、それに伴う特殊な構造(caudalrib の有無など、挿図5)が限られたグループにみられることから、後方への延長は本亜目内の類縁関係を推定する上で有効な形質となり得ると判断された。すなわち、いくつかのグループにおいては、後方への延長がそれぞれのグループの単系統性(共通の祖先をもつこと)を支持する形質(共有派生形質)となり得ると考えられた。
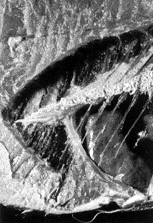
 | 29-3 イケカツオ Scomberoideslysan(アジ科)。 ZUMT57763、体長170mm、日本。 |
 |  |
| 29-5 Schuetteasclaripinnis (ヒメツバメウオ科)。ZUMT55205、体長105mm、オーストラリア。 | 29-6 Chaetodipterusfaber (スダレダイ科)。ZUMT57747、体長95mm、スリナム沖。 |
 |  |
| 29-4 タテジマキンチャクダイ Pomacanthusimperator(キンチャクダイ科)。ZUMT52830、体長97mm、沖縄。 | 29-7 スダレダイ Drepanelongimana(スダレダイ科)。HUMZ46947(註17)、体長138mm、ボルネオ沖。 |
しかしながら、延長部の構造が多様性に富むことから(口絵、挿図3〜7参照)、2つ以上のグループにみられるそれぞれの延長部が相同であるかどうかを判断することは極めて困難であり、多くの場合、相同であるとは考えられなかった。このことは、後方への延長という現象が、スズキ亜目魚類の進化の過程において、いくつかのグループで独立に何度も生じたことを強く示唆している。 スズキ亜目魚類の類縁関係を推定する上で、鰾の後方延長部にみられる多様性の進化の解明が今後の課題の1つとなるが、これには遊泳、摂餌などの行動および個体発生からの情報が不可欠であると考えている。
(坂本一男)
註1 富永義昭、1991. 総合研究資料館ニュース、(22)、1〜3 坂本一男、1994、総合研究資料館ニュース、(22)、6〜7