二枚貝類(Bivalvia)
- 貝殻
- 貝殻は体の左右に位置します。二枚貝類の2枚の殻は、単板類型の1枚の殻が折れ曲がってできたと考えられています。絶滅した軟体動物の吻殻類(Rostroconchia)には、笠型の貝が左右に押しつぶされたような殻をもつ種があり、これが単板類型貝類から二枚貝類への移行段階を表していると解釈する説があります。左右の殻はそれぞれ殻片(valve)と呼ばれます。二枚貝類には無殻のものはありません。ただし、渦虫状のXenotur bellaは、幼生が原鰓類の幼生に類似するため無殻の二枚貝類である可能性が指摘されています。
二枚貝の殻の向きは、足があり殻が開く方が腹縁(ventral margin)、咬み合わさっている方が背縁(dorsal margin)、軟体の口がある方が前縁(anterior margin)、水管側が後縁(posterior margin)です。殻の成長の原点は殻頂(umbo=beak)と呼ばれます。殻頂には幼生期の貝殻が残されており、原殻(prodissoconch)と呼ばれます。殻頂の位置は中央にある場合(equilateral)と、前後に偏る場合(inequilateral)があります。殻頂の傾きには前傾(prosogyrate)と後傾(opisthogyrate)が区別されます。
左右の殻には、等しい形態の等殻(equivalve)と、左右不相称の不等殻(inequivalve)があります。不等殻は固着性の種に多く見られます。
2枚の殻は背側で弾力性のある靭帯(ligament)によって結びつけられています。殻は閉殻筋(adductor muscle)が収縮することによって閉じられ、閉殻筋が弛緩すると靭帯の力で開きます。靭帯は殻の殻頂の外側にある外靭帯(ligament)と殻頂の内側にある内靭帯(resilium、複数形resilia)に区別されます。内靭帯は靭帯受(resilifer)に付着し、靭帯受が突出している場合は弾帯受(chondrophore)と呼ばれます。靭帯は石灰質の針状結晶をほとんど含まない外側の層状靭帯(lamellar layer)と、針状結晶を含む内側の繊維状靭帯(fibrous layer)の二層からなります。
外靭帯の形式にはマルスダレガイ科Veneridaeなどに見られる筒状靭帯(parivincularligament)、シラスナガイ科Limopsidae、ウグイスガイ科Pteriidae、イタボガキ科Ostreoideaなどに見られる単純靭帯(alivincular ligament)、フネガイ類Arcoidaなどに見られる重複靭帯(duplivincular ligament)、マクガイ科Isognomonidaeなどに見られる多靭帯(multivincular ligament)などの形式があります。
殻の背中側の接線は 線(hinge line)と呼ばれます。線の内側はやや厚くなり板(hinge plate)が形成され、その上に左右の殻を咬み合わせるための歯(hinge teeth)が発達します。歯には、多歯型(taxodont)(図1-15)、貧歯型(dysodont)、等歯型(isodont)、裂歯型(schizodont)(図1-16)、異歯型(heterodont)などの形式があります。異歯型の場合、主歯(cardinal teeth)と側歯(lateral teeth)が区別され、後者は前後の位置によって前側歯(anterior lateral teeth)あるいは後側歯(posterior lateral teeth)と呼ばれます。 線(hinge line)と呼ばれます。線の内側はやや厚くなり板(hinge plate)が形成され、その上に左右の殻を咬み合わせるための歯(hinge teeth)が発達します。歯には、多歯型(taxodont)(図1-15)、貧歯型(dysodont)、等歯型(isodont)、裂歯型(schizodont)(図1-16)、異歯型(heterodont)などの形式があります。異歯型の場合、主歯(cardinal teeth)と側歯(lateral teeth)が区別され、後者は前後の位置によって前側歯(anterior lateral teeth)あるいは後側歯(posterior lateral teeth)と呼ばれます。


- 軟体部外部形態
- 体は左右に偏圧されます。足は斧状で足裏をもつものはごく少数です。頭部は分化していません。
外套膜は殻と靭帯を形成し、内臓塊を覆います。外套膜は細い筋肉で殻に付着しており、その付着部が殻表上に残る跡痕のことを外套線(pallial line)と呼びます。外套膜は左右二葉に分かれていますが、しばしば後ろ側で癒合して、入水管(inhalant siphon)と出水管(exhalant siphon)を形成します。水の取り入れられる経路は決まっており、腹側が入水管、背側が出水管です。この向きは肛門が背側に開口することと関係しています。
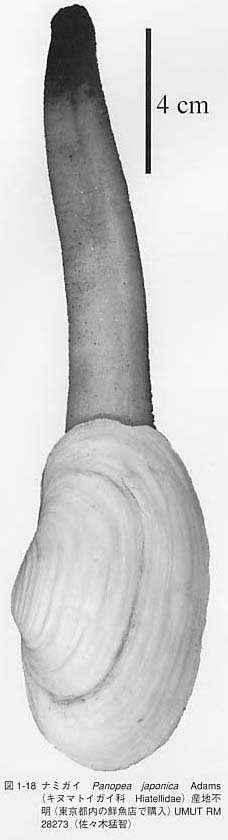
水管の納まる部分の外套線は湾入しており、外套湾入(pallial sinus)と呼ばれます。外套湾入の深さは水管の長さにほぼ比例し、水管が長い種ほど底質中に深く潜入する能力を持ちます。従って、外套湾入の深さから水管の長さと二枚貝が潜入する深さを推定することができます。水管はほとんどの種では殻の中に退縮させることができますが、ミルクイTresus keenae(図1-17)、ナミガイPnopea japonica(図1-18)、ウミタケガイBarnea(Umitakea)dilatataなどは水管が殻に収納できないほど長い水管を持ちます。
外套膜縁の断面は外褶(outer fold)、中褶(middle fold)、内褶(inner fold)の3葉に分かれます。内褶は最も厚く、放射状の筋肉を含みます。中褶には外套触手(pallial tentacle)が発達して感覚器としての機能を持ち、外褶は貝殻の形成に関与しています。
二枚貝類には頭部眼がありません。しかし、イタヤガイ科Pectinidae、ウミギクガイ科Spondylidaeでは外套膜の中褶に外套眼を発達させます。一方、ザルガイ科Cardiidaeでは水管の感覚触手に光受容器官を持っています。
多くの二枚貝は前後に閉殻筋を持っており、双筋型(dimyarian)と呼ばれます。しかし、固着性の二枚貝では足と前閉殻筋が退化的になり、不等筋(anisomyarian)と呼ばれます。さらには、イタヤガイ科のように前閉殻筋が完全に失われるものがあり、単筋型(monomyarian)と呼ばれます。足を引っ張る収足筋(pedal retractor muscIe)は体の前後にあります。前閉殻筋と後閉殻筋のすぐ内側で殻に付着し、独自の付着痕を殻の内側に残します。
二枚貝には足糸(byssus、複数形byssi)と呼ばれる糸を使って貝自身を海底の基質に固定するものがあります。この足糸は二枚貝に特有の付着器官です。足糸は足にある足糸腺(byssal gland)で分泌されます。足糸腺の末端からは足の表面に溝が走っており、足糸の元になる液が分泌されると足の溝の中を流れて付着面に到達します。液体は海水に触れると硬くなり糸状になります。足糸の牽引筋を収縮させれば貝は付着面にぴったりと固着することができます。足糸の形成は二枚貝にとって原始的な形質と考えられます。発生過程では幼生は足糸腺を持ちますが、多くの二枚貝が成長にともなって足糸を失います。セメント質で固着する種の幼生も、まず足糸を出して体を固定したのちに殻を固着させます。生息環境との位置関係では、基質の表面に足糸で付着する様式をexobyssate、底質中に潜り礫などに付着する様式をendobyssateといいます。
- 外套腔
- 二枚貝類の外套腔には大きな鰓が含まれます。鰓は鰓葉の形と鰓葉間の連絡の形式により、原鰓型(protobranch)、糸鰓型(filibranch)、弁鰓型(lamellibranch)、隔鰓型(septibranch)の4型が区別されます。異歯類Heterodontaは全て弁鰓型を持ちますが、翼形類には糸鰓型と弁鰓型の両者が見られます。系統学的に両者の弁鰓型を別起源と考える場合には、前者を真弁鰓型(eulamellibranch)、後者を擬弁鰓型(pseudolamellibranch)と見なします。
二枚貝の鰓は外套腔内に左右に一つずつあります。鰓には血管系と繊毛が発達します。それぞれの鰓の鰓軸の両側には鰓葉(ctenidial lamella、複数形lamellae)が発達し、外側を外半鰓(outer demibranch)、体の中心側を内半鰓(inner demibranch)と呼びます。
原鰓型では、2つの半鰓は小さく、他の軟体動物と同様の原始的な二葉型の鰓を持ちます。
糸鰓型ではそれぞれの鰓葉は糸状の鰓糸(ctenidial filament)からなります。鰓糸には網目状の隙間があり、組織の結合部は鰓葉結合(interlamellar junction)によって連結しています。隣接する鰓系は繊毛によって支えられます。多くのグループでは鰓糸が背側に折り返し、上行枝(asending arm)と下行枝(descendin garm)が区別されます。繊毛によって捕らえられた粒子は、外半鰓の上行枝の基部と内半鰓の下行枝の末端にある食溝(food groove)に集められます。
弁鰓型は糸鰓型に類似しますが、鰓葉は鰓葉結合に加えて鰓糸結合(inter filamental junction)によって結びつけられ、格子状になります。その間は長い柱状の空間になりwater tubeと呼ばれます。鰓糸結合の間には小さな孔(ostium、複数形ostia)があり、水流はこの孔を通じてwater tubeに入り込み、背側に上がって出水管へ排出されます。
隔鰓型では鰓は変形して隔膜(septum)になり、濾過食の機能は失われます。
化学的受容器官である嗅検器は後閉殻筋の前側の鰓の基部にあります。二枚貝の嗅検器は他の軟体動物の嗅検器よりもかなりわかりにくい形をしていますが、色素が含まれている場合には外表面からも識別できます。嗅検器は脳神経節に由来する神経節に支配を受けます。二枚貝の嗅検器は水が流入する側ではなく、流出する側にあります。一方、腹足類では逆に位置しています。それゆえ腹足類の嗅検器との相同性はよくわかりません。
- 消化器官
- 二枚貝類には頭部がなく、単純に口が開くのみです。口の前側には1〜2対の唇弁(labial palp)があり、鰓から送られてきた餌粒子を選別し、口へ運びます。大型の粒子は排除され、殻外へ排出されます。
全ての二枚貝類には歯舌も口球もありません。消化管の前部は単純で、唾液腺や食道腺は発達しません。胃には1対の消化腺が付属します。胃の内部には消化酵素を含むすりこぎ状の晶体=晶桿体(crystalline style)が発達します。腸は長く、足の中で何度も螺旋を描き、肛門は出水管の基部付近に開口します。
原鰓類は懸濁物食者ではなく堆積物食者(deposit feeder)です。2対の唇弁状の構造を持っていますが、外側の唇弁は唇吻(palp proboscis)と呼ばれ、殻の外側まで伸ばすことができます。そして、デトリタスを表面の粘液によって捕らえ、繊毛によって口へ運びます。内側の唇弁は他の二枚貝の唇弁と同様です。
弁鰓類では、鰓が摂餌のために利用されます。水は流入する側から流出する側へ鰓の側方繊毛(lateral cilia)によって運ばれます。水が鰓糸間を流れるときには正面側方繊毛(laterofrontal cilia)の間を通り抜けます。これらの繊毛は粒子を捕らえるのに役立ちます。鰭糸の表面では粒子は正面繊毛(frontal cilia)の働きによって、鰓の末端にある食溝へ、そして唇弁へと運ばれます。拒絶された粒子は外套腔へこぼれ落ちて、擬糞(pseudofeces)として排出されます。
隔鰓類は肉食性の捕食者です。隔鰓類の外套腔は鰓が変形してできた隔壁によって背側と腹側の空所に分けられます。筋肉の動きにより、隔壁は外套腔の中で上下に動きます。隔壁を上に上げると入水管から水が吸入され、隔壁を下げると水が背中側へ移動して、出水管から排出されます。この動きを規則的に繰り返すことにより、小型の甲殻類を外套腔へ吸い込み、筋肉質の唇弁によって捕らえて捕食します。
隔鰓類の胃は著しく角質化されており、餌を細かく砕く器官として機能します。他の二枚貝類にみられるような晶体は退化的になります。キヌタレガイ類Solemyaは鰓に共生細菌をもち消化管が退化します。化学合成生物群集に棲息する二枚貝類には消化管が単純化する傾向がみられます。
- 排出器官
- 二枚貝では排出器官は左右1対でU字状に曲がっています。U字状の腕の一方は腺状部で、囲心嚢に開きます。もう一方の腕は膀胱(bladder)を形成し、鰓上腔(suprabranchial cavity)と呼ばれる鰓の後部上方の空間に開きます。
- 循環器官
- 二枚貝の心臓は2心房1心室です。多くの二枚貝では心臓は腸によって貫かれています。しかし、いくつかの分類群では腸は心臓を迂回します。例えば、カキ上科Ostreoideaのベッコウガキ科Gryphaeidaeでは腸が心臓の中を貫通しますが、イタボガキ科Ostreidaeでは腸は心臓の外側を通ります。
- 生殖器官
- 二枚貝の生殖器官は非常に単純で、生殖輸管は外套腔に直接開口するか、腎臓を介して外套腔に開口します。二枚貝類はほとんどの種は雌雄異体ですが、性転換をするものも知られています。いくつかの雌雄同体の種では、単一の生殖巣が卵と精子の両者をつくり、単一の生殖輸管で開きます。いくつかの例では卵巣と精巣が分離して、生殖輸管もその開口部も別々につくるものがあり、それらの中間的な構造を持つものも知られています。
- 神経系
- 二枚貝の神経系は主に脳神経節、側神経節、足神経節からなります。原鰓類では脳神経節と側神経節が部分的に癒合します。その他の二枚貝では、脳神経節と側神経節が完全に癒合して1つの脳側神経節(cerebropleural ganglion)になります。そのため脳神経節と側神経節の縦連合(cerebropleural connective)がありません。脳側神経節は内臓神経節と足神経節に向かって神経を伸ばし、唇弁、前閉殻筋、外套膜に神経を送ります。口球が無いため口球神経節は見られません。内臓神経は消化管、心臓、鰓、外套腔、水管、後閉殻筋に神経を送ります。
|