頭足類(Cephalopoda)
- 貝殻
- 頭足類には貝殻をもつ種ともたない種があります。オウムガイ類は外殻性の殻を持ち、内部は隔壁(septum、複数形septa)によって仕切られています。隔壁をもつ部分は気房(air chamber)=房錐(phragmocone)、隔壁の内部は気室(chamber = camera)と呼ばれます。隔壁の内部は連室細管(siphunclular tube)によって貫かれ、その内部には体管索(siphuncular cord)と呼ばれる組織がつまっており、浮力調節器官として機能しています。動物体は殻の末端部にある住房(body chamber = living chamber)に入っており、動物体は成長するにつれて定期的に前方へ移動し、後方に新しい隔壁をつくります。現生オウムガイ類の場合隔壁は30以上形成されます。殻の殻頂部には胚の段階で形成される胚殻(embryonic shen)が残されています。
頭足類の貝殻は外殻性から内殻性に進化したと考えられています。現生種では内殻性が一般的です。トグロコウイカSpirula spirula(図1-10)ではオウムガイ類と同様の螺旋状に巻いた殻を持っていますが、殻は内殻性で大部分が外套膜によって包まれています。
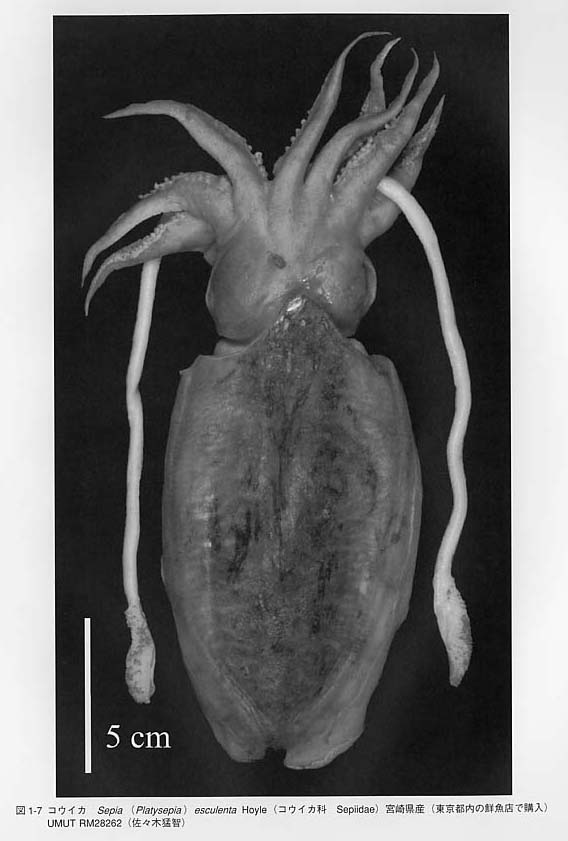
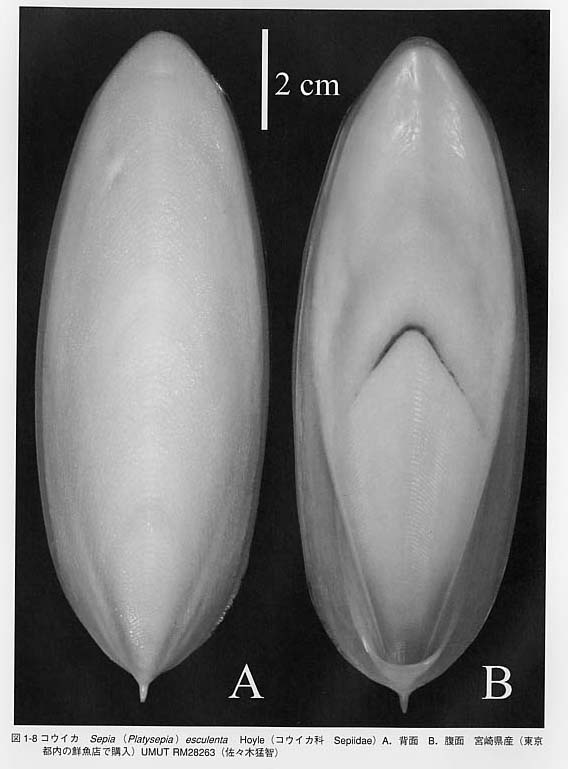
内殻性の殻はさらに退化したり、消失します。コウイカ類Sepiaの貝殻は背腹に押しつぶされたような形をしています(図1-8)。オウムガイ類やトグロコウイカとは無関係の形態に見えますが、断面を見ますと、内部には薄い板状の隔壁が密集しており、その間には隙間があります。この空間はオウムガイ類やトグロコウイカの気室に相当します。石灰質の殻はツツイカ類では有機質の膜になり軟甲(gladius = pen)と呼ばれます。さらにタコ類の成体では貝殻は形成されません。
アオイガイ類・タコブネ類Argonauta(図1-11)も螺旋状に巻いた外殻性の貝殻をもちます。しかし、この殻の内部には隔壁がありません。この殻は卵を保護するために雌の第1腕から分泌されたもので、雄にはありません。他の頭足類の殻は腕ではなく外套膜から分泌されるため、両者の殻は起源が異なります。
- 外部形態
- 頭足類の体は前後方向に長くなり、腹側に深い外套腔が形成されます。体は左右対称です。腕(arm)と口がある方が前方で、内臓塊は体の後方にあります。 漏斗の周辺の頭部の最後端部は頸部(neck)と呼ばれます。漏斗の左右には頸部と外套膜を咬み合わせる漏斗軟骨器(funnel locking cartilage)と呼ばれる軟骨の塊があります。
ツツイカ類では体の後方に鰭(fin)があります。コウイカ科Sepiidaeでは外套膜の側方全長にわたって鰭が延長しています。アオリイカSepioleuthis lessonianaも同様の鰭をもち紛らわしいですが、コウイカ類ではなくツツイカ類のヤリイカ科Loliginidaeです。タコ類には鰭の無い種が主流ですが、有触毛類Cirrataには全て鰭があります。コウモリダコVampyroteuthis infernalisにも同様の鰭があります。ツツイカ類の鰭は体の向きを安定させる機能があります。コウイカ類では鰭を波打たせており、遊泳に役立っているようです。
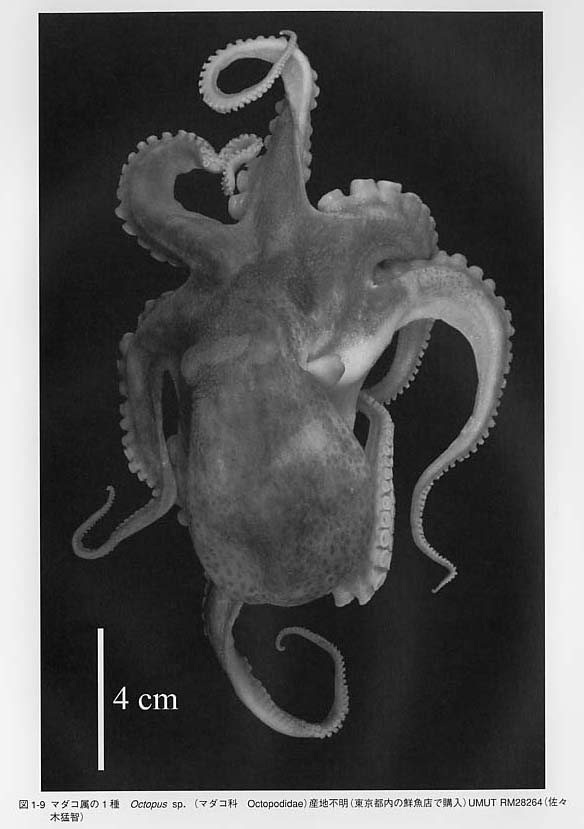
腕は口を取り囲むように分布しており、背中側から左右にI、II、III、IVと数えます。コウイカ類・ツツイカ類では、III腕とIV腕の間から伸びる長い腕は触腕(tentacle)と呼びます。この2本の触腕の有無により「タコは8本、イカは10本」の差を生じます。種によっては腕の間に傘膜または腕間膜(umbellella = interbrachial membrane)と呼ばれる広い膜が発達するものもあります。雄では一部の腕に生殖のための特殊化が見られます。
腕はタコ類は8本で、イカ類は10本です。ところが、これには例外があります。ヤツデイカ科Octopoteuthidaeに属する種とテカギイカ科GonatidaeのタコイカGonatopsis borealisはイカ類でありながら成体では腕が8本しかありません。幼若個体には10本腕があるのですが、成長に伴って触腕が2本とも失われてしまいます。オウムガイ類では腕は
全部で90本近くあります。
鞘形類の腕には吸盤(sucker)や鈎(hook)を持っています。オウムガイ類の触手には吸盤がなく、粘着性の分泌物で付着します。イカ類の吸盤は基部が柄のように細くなっており、吸盤の内部には角質環(chitinous ring)と呼ばれる硬い有機質のリングがあります。一方、タコ類には柄も角質環もありません。この吸盤の構造の違いが「イカ」と「タコ」を区別する最も重要な形質です。メンダコ類の腕には吸盤に加えて触毛(cirrus、複数形cirri)が生えています。タコ類の吸盤は吸着によって、イカ類の吸盤は角質環でしがみつくことによって機能するという違いが見られます。
外套腔の入り口付近の外套膜の縁辺部は襟(collar)と呼ばれます。イカ類やオウムガイ類では外套膜は体の外側を取り囲んでおり、外套腔の出入り口は背側にも腹側にも存在します。一方、タコ類の外套膜は背中側で頭部の表面と癒合しており、両者の境界線が見られません。
頭足類は優れた運動性能を持ちます。腹側には漏斗(funnel)があり、そこから水を吹き出して、ジェット推進(jet propulsion)で移動することができます。頭足類の外套膜には環状筋と放射筋が重なり合っており、環状筋がゆるみ放射筋が収縮すると頭と外套膜の間から水が取り入れられます。次に環状筋が収縮すると襟が頭部に押しつけられ、水は漏斗から排出されます。漏斗の向きは自在に動かすことができ、これによって遊泳方向を調節できます。漏斗はオウムガイ類では閉じた管ではなく、それ以外の頭足類では完全に閉じた水管状です。
頭部には眼がよく発達しています。オウムガイ類のみはレンズのないピンホール型の眼を持ちます。鞘形類はレンズを伴う高度に発達した眼を持ちます。コウイカ類では眼の表面は隔膜で被われず開いています。ツツイカ類ではヤリイカ科Loliginidaeとピックフォードイカ科Pickfordiateuthidaeが閉じた眼を持っています。そのため、ヤリイカ類を閉眼類Myopsida、それ以外のツツイカ類を開眼類Oegopsidaと呼びます。
頭足類の眼は脊椎動物の眼に酷似します。しかし、頭足類では視神経が網膜の外側から
伸びるため盲点がありませんが、脊椎動物の眼では内側から伸びた視神経が束になって網膜を貫通する点に盲点が形成されます。
- 消化器官
- 頭足類は背腹に1対になった強力な顎板を持ちます。頭足類の顎板は一般には「カラストンビ」と呼ばれています。顎板は黒い有機質の板ですが、オウムガイ類では先端部が厚く白い石灰質に被われ、そこに歯のような構造を持ちます。頭足類の歯舌はオウムガイ類では13本(中歯+側歯2対+縁歯2対+縁板2対)ですが、アンモナイト類では9本(中歯+側歯2対+縁歯1対+縁板1対)、鞘形類では9本または稀に7本です。ただし、トグロコウイカには歯舌がありません。大部分の頭足類は口球の後側に2対の唾液腺を持ちます。後方の唾液腺は毒線として機能しており、毒で餌を麻癖させます。しかし、オウムガイ類は前側の1対の唾液腺しか持ちません。
オウムガイ類以外の頭足類は危険を感じると墨を吐き出します。墨は直腸の付属腺である墨汁嚢(ink sac)に蓄えられており、漏斗から吐き出されます。墨はタコ類では拡散
して煙幕として機能します。イカ類では墨は拡散しにくく、捕食者の眼を墨の塊にそらす働きがあります。深海性の頭足類には発光性の光る墨を吐く種があります。
頭足類は肉食性の捕食者です。遊泳性のイカ類は魚類、底生性のタコ類はカニや二枚貝を好んで捕食します。タコ類は貝類に対しては歯舌で孔を開けることができます。タコ類では吸盤に化学受容細胞が分布しており、視覚に頼ることなく触覚で餌を採るともできます。例えば、テナガダコOctopus minorは海底の泥の中に埋もれたまま、第1腕のみをのばして餌を捕獲します。
- 循環器官
- 循環器系はほぼ閉鎖血管系です。鰓はオウムガイ類が2対、それ以外の現生種は1対です。かつては鰓の数に基づいて二鰓亜綱Dibranchiataと四鰓亜綱Tetrabranchiataに分ける分類体系が用いられていましたが、絶滅種には適用できないため現在では用いられていません。心臓の心房の数も鰓の数に対応しており、オウムガイ類では2対、オウムガイ類以外の頭足類では1対です。鯉の基部には鰓に効果的に血流を送るための鰓心臓(branchial heart)が発達します。これは鰓に入る血管の一部が膨出してできたものです。
- 排出器官
- オウムガイ類のみが2対、それ以外の頭足類は1対の排出器官を持ちます。排出器官は腎囲心嚢連絡管によって囲心嚢と連絡しており、囲心嚢で濾過された排出物質が排出器官に送られます。排出器官の一部は膨れて腎嚢(renal sac)を形成します。近くを通る大静脈(vena cava)が袋状に突出し、囲心嚢側には囲心嚢付属腺(pericardial appendage)、腎嚢側には腎嚢付属腺(renal appendage)と呼ばれる膨らみを形成します。これらの付属器は規則的に収縮し、排泄物質を排出器官に送る機能を持っています。
- 生殖器官
- 全ての頭足類は雌雄異体です。体の後部に単一の生殖巣があり、生殖輸管が前方へ伸びます。
雄の輸精管は1本しかなく、オウムガイ類では右側、鞘形類では左側にあります。雌の輸卵管は、開眼類の大部分とタコ類のうち無触毛類Incirrataでは2本ありますが、オウムガイ類では右側のみ、コウイカ類と閉眼類とタコ類のうち有触毛類Cirrataでは左側にしかありません。輸卵管の末端部には輸卵管腺(oviducal gland)が付属しており、卵殻を分泌します。輸卵管の開口部の近くには包卵腺(nidamental gland)があり、卵を保護するための物質、あるいは卵を地物に付着させるための物質を分泌します。
精子は輸精管の一部が変形してできた貯精嚢に蓄えられます。精子は精包(spermatophore)に入れられて雌に渡されます。精包は前立腺(prostate gland)からの分泌物が貯精嚢に入り込み、精子を固めることによって形成されます。完成した精包は輸精管の末端部の膨らんだ部分、ニーダム嚢(Needham's sac)に交接直前まで蓄えられます。精包は外套腔から交接腕(hectocotylus)を通じて雌に渡されます。
雌の輸卵管の開口部は外套腔の奥にあります。そのため、雄は交接腕を使って、雌の外套腔に精包を送り込みますが、イカでは雌の特定の部位、例えば囲口膜、口唇、襟、外套膜などに精子塊が植えつけられます。交接腕には特別な吸盤や、精包をつかむための構造が発達します。イカ類では右または左、あるいは両方の第IV腕が使われます。タコ類では右の第III腕です。オウムガイ類では交接用の腕は円錐形の器官に変形しておりspadixと呼ばれます。
アオイガイ類Argonautaでは雄の腕は交接中にちぎれて、雌の外套腔内に残ります。キュビエはそれを寄生虫と考え、他Hectocotylus octopodisと名付けました。一方、さらにケリカーという頭足類学者は、残された腕を完全な雄個体と信じ込み、心臓や消化管や生殖器官の存在までを記載したという逸話も残っています。
鞘形類では一般的に雄よりも雌が大きくなります。例えば、浮遊性のタコ、ムラサキダコTremoctopus violaceusは雌は全長50cmを越えますが、雄は3cmくらいです。逆にオウムガイ類、ヤリイカ類では雄が大きくなります。
- 神経系
- 神経系の発達と集中化は頭足類の特徴です。鞘形類では、脳神経節、足神経節、側神経節、内臓神経節が全て癒合し、食道の周囲にリング状の神経環を形成し、脳(brain)と呼ばれます。そして、脳は軟骨質の頭蓋(cranium)に取り囲まれています。このように中枢神経系が軟骨に包まれる構造は他の軟体動物には見られません。脳の塊には瘤状の部分が識別でき、葉(lobe)と呼ばれます。
例えば、タコ類では頭頂葉、上位前額葉、下位前額葉、上位口球葉、視柄下葉、内臓葉、外套葉、後部色素胞葉、前部色素胞葉などの名前が付けられています。脳の前方の腕にはタコ類では4対、イカ類では5対の腕神経(brachial nerve)が伸びます。外套腔の背中側の外套膜には巨大な星状神経節(stellate ganglion)が発達していますが、この神経節は他の軟体動物には見られません。頭足類は視覚・触覚・遊泳行動の能力に優れ、学習する能力を持つことが知られていますが、それは脳の発達と関係しています。平衡胞は、オウムガイ類では他の軟体動物と同様に足神経節の近くにありますが、鞘形類では頭部軟骨の腹部にある平衡胞中に存在しています。
|