自然界の贋作
|
高槻成紀 総合研究博物館・動物生態学 井上泰輔 文学部・美術史学 阿部修英 文学部・美術史学 高堂貴 文学部・考古学 |
|
本企画全体のテーマは真と偽について考えることにある。私たちはそのうち動植物にみられる真と擬、つまり擬態について分担することにした。初めに擬態についての概論を述べ、それから実例を紹介し、最後に自然界の贋作ともいる擬態の意義について考えてみたい。 初めに語句について整理しておきたい。擬態は「擬」つまりだまされるほどよく似ているということと「態」つまり動物の形態とか生き方などの状態の二字から構成されており、他の動物を形や動きで騙すことをいう。少なくとも日本語の擬態という語からはそういうことがイメージされる。 英語で擬態を意味するmimicryはギリシア語のミモス(mimos)、つまり演技とか模倣を意味することばに由来し、パントマイムのマイム(mime)にも派生した。これが生物学の専門用語となるのだが、日本語の擬態のほうが優れているように思う。 ところで生物を離れて一般に、偽りにしても贋作にしても、コピーされる側の本物(モデル)があることが前提となる。これがなければそれは創作になってしまう。コピーにはふたとおりあり、その違いは本稿では本質的なものとなる。ひとつはコピーであることが見るものにとってマイナスにならないものである。たとえば、双児は互いによく似ているが、そのことは親にとっても周囲の人にとってもなんらマイナスではない。何々モドキという似て非なる動植物があるが、これらも同様である。ウメモドキはあまりウメに似ていると思えないが、ウメに似た植物という意味である。ツルウメモドキはツル植物でウメとは似てもつかないが一枚の葉をみると確かにウメの葉に似ている。「モドキ」とはいわないがエノキグサという植物があり、この葉はエノキにそっくりだ[写真1]。ゲンゴロウは子供に人気のある水生昆虫だが、これに似た「シャープゲンゴロウモドキ」という種がいる。日本列島で進化した種であるのに「シャープ」という形容をつけられ、しかも「モドキ」とされたのではずいぶん失礼な気がする。しかし「モドキ」はまだよいほうかもしれない。「ゴミムシダマシ」、「ニセリンゴカミキリ」、「ニセヨゴレイタチシダ」などという気の毒なような名前をつけられた動植物もいるのだから。
しかし、これらはみな人間が似ていると感じるだけのことで、ウメやゲンゴロウがもともとあって、あとから「モドキ」が現れたわけではもちろんない。人間には基本的な認識があり、それを拡大するときに形容詞をつけて区別しながら認識を拡大してゆく。チゴユリ、ヒメサユリ、ウバユリとか、アサガオとヒルガオのような場合は中立的だが、チョウセンアサガオは朝鮮から来た朝顔ということであり、日本人には朝顔が「あるべきもの」で、チョウセンアサガオは外来者ということになる。先にあげたような「モドキ」や「ダマシ」の場合も同様で、主たるものがあり、それに似たものという認識のヒエラルキーがある。見たことがないものに出会うと、それまで知っていたものに形容詞をつけることになる。エゾマツは蝦夷の松であり、アメリカザリガニはアメリカから来たザリガニである。タヌキはまぎれもなくひとつの種だが、タヌキのいない英語圏ではracoon-dogつまり、アライグマのようなイヌ(アライグマモドキ?)と呼ばれる。そもそもアライグマは「クマ」ではなく、日本にいないこの動物を自分たちの知っている動物のなかではクマに似ているものに「物を洗う」という形容をしたものである。もしタヌキモドキとかアライダヌキとしていたら、英語と日本語がお互いにないものをそれまでに知っていたものの「モドキ」とした互換性の好例となっていたと思うのだが。 さて、コピーのもうひとつは似ていることが見るものにとってマイナスとなるもので、これが偽物とか贋作とか呼ばれる。ニセモノのニセは「贋」と書くが、大和ことばでは「似もの」に違いなく、「似たもの」ではない。似たものが前述のマイナスでないコピーであり、ことばは単に似ているという状態を示す。これに対して「似もの」は似たという意図、それも邪(よこしま)な意図が感じられる。本物に価値があり、コピーには価値がない、それを見た者がまちがえて騙されることによって被害が生じると、そのコピーは贋作になる。偽札や偽宝石、そして絵画や彫刻などの芸術作品に典型的にみることができる。 こう考えると美術作品の中心的存在である自然や人物、静物をオブジェとした作品がまちがいなくそれらのコピーであり、芸術行為そのものがコピーしたいという欲求の表出なのではないかという気がしてくる。これらの場合、オブジェがまさに本物であって、作品とは本質的に異なるものであることである。これに対して、いわゆる贋作はその「本物の」作品のコピーであり、たとえば絵画であればカンヴァスの上に絵具がのっているという意味では物質としては同質なものである。この違いは本質的である。 以上のように贋作に関する語句と概念を整理した上で本題である擬態を考えてみたい。擬態の代表例であるハチとこれに似た昆虫群を考えてみよう。ハチは刺されると激痛をともなう危険な昆虫である。そのような外敵を刺す危険な昆虫でとくに目立たない種がいたとする。そのハチに触れた哺乳類や鳥類が刺されて痛い目にあうと、そのことは記憶される。もう一度あったときはハチに触れないようにするだろう。このときそのハチが記憶に残りやすいほうが、再発防止が効果的になるはずである。このハチのなかから遺伝的変異によって多少色の変わったタイプが生じたとしよう。黒や茶の目立たないものよりも黄と黒の縞模様のある個体のほうが「私はあの危ないハチだぞ」というアピールが効果的になり、そのような個体の生存率が高まるだろう。このような繰り返しによって現在のスズメバチに代表されるような目立つ体色のハチが生じたと考えられる。 このようなモデルが生じれば、そのコピーが生まれるのは必定である。アブは無害であるが、ハチに大変よく似ているので、哺乳類や鳥類はハチと同じように反応する。トラフカミキリはコガネムシと同じ甲虫(鞘翅目)に属し、ハチとは系統的に遠く離れているが、その形態がハチに似ていることは驚くばかりである。このような「贋バチ」が、ハチで痛い目にあった哺乳類や鳥類の捕食をまぬがれて利益を得ていることはまちがいない。これらのグループにおいても地味な色の個体のほうが捕食率が高くて集団内で「ハチ型」が増加したものと考えられる。このような進化は「騙される」捕食者がいることが前提となる。そうでなければハチをまねることは意味のない独り芝居にすぎない。 それではこの「贋バチ」は騙されるものにとってマイナスだろうか。捕食者にとって食物資源となりうるものがこけおどしのために食べることができなくなっているとすれば、これはマイナスとなっているといるだろう。その意味でこのコピーは贋作といる。 食べられることを回避することは死亡要因にかかわるから進化の選択圧となる。ほかの動物をコピーするのを狭義の擬態というが、食べられることを回避するという意味ではまわりの景色に溶け込んで発見されにくくするカモフラージュ(迷彩)も同質の機能をもっている。だいたいにおいて褐色系の色は土や枯葉、枝や幹など自然界に多い色なので迷彩効果がある。蛾の仲間には褐色系の翅のものがいるが、樹木の幹にとまっていると完全に見失うほど迷彩効果がある。ヒキガエルなどの体色も地面の色や質感とよく似ている。逆に青々とした緑のなかでは緑色に迷彩効果があるからバッタの仲間にはイネ科の植物と実はよく似た質感の体色をもつものがいる。きわめつきは、その名もコノハムシという直翅目の一種であろう[写真2]。体全体が一枚の葉に似ているとうだけでなく、6本の脚のすべてが葉のようだし、その質感たるや緑の枯れかかった感じとか、表面の汚れまで実に念が入っている。
水中でも同様の現象がみられ、海藻にそっくりなタツノオトシゴの仲間のリーフィー・ドラゴンフィッシュ(葉のようなタツノオトシゴ)や、海底の砂に溶け込んでしまうヒラメやカレイなど枚挙にいとまがない。 迷彩効果は見えないことを旨とするのだが、これを逆手にとるダミー効果もある。タテハチョウ類によくみられる翅の裏面の褐色は蛾の翅の表面同様、まわりの景色に溶け込んでしまうが、この場合、捕食者にとっては表面の鮮やかな色彩が印象的なために、翅を閉じたときの地味な色との落差が大きく捕食者に「消えた」と感じさせる効果がある。飛翔性バッタの仲間には翅を開いたときに内側に鮮やかな紅色系の翅をもつものがいて、飛んでいるときはこの色が目立つが、着地して翅を閉じると突然その色が消えるので、草の緑のなかに「消える」ことになる。このような視覚の落差を使うのはかなり高度な迷彩効果といるだろう。これは追跡者に対象物を「延長」する感覚が働くためである。追跡者は逃走者の次の動きを読もうとする。刑事が犯人を追跡する場合、角を曲がったらその先の動きを予測する。賢明な逃走者は走路を予想外のものにしたりして「巻こう」とする。実際にあったことらしいが、逃走者が角を曲がったあとでビルに入って女装し、堂々と逆方向に歩いて追跡を巻いた事例があるという。筆者(高槻)は野外でサルを追跡していて、尾根を越えたサルがそれまでと歩く速さとコースを完全に変えたためにみごとに巻かれた経験がある。これは形態や色彩の特殊化をともなわない逃走法であるが、それでもこれだけの効果があるのだから、これらがともなう逃走には大きな迷彩効果があるに違いない。 以上、食べられること回避するための擬態を考えたが、擬態にはこれ以外にもさまざまなものがある。同じ迷彩効果でも目的が捕食である例もある。自然界では食べる、食べられる関係をめぐってさまざまな工夫が見られるから、食べるほうが効率を高めるために採用される工夫のなかに迷彩効果があるのは当然といる。クモ類は花に訪れる昆虫を捕らえるために自分を目立たなくして花の近くに潜むので迷彩をする。迷彩というより隠蔽というべきかもしれない。東南アジアにいるハナカマキリはカマキリとは思えぬピンク色をしており、花にそっくりである[写真3]。これほどに見事なコピーに驚きを感じない人はいないだろう。
次に相手を騙すの例として有名なカッコウ類の托卵をとりあげてみよう。 カッコウ類は初夏に温帯に渡ってくる渡り鳥で,その印象的な鳴き声によってよく知られている鳥だ。温帯に渡ってくるのは繁殖のためだが、かれらは実は変わった繁殖をする。巣を作らないばかりか、自分の産む卵をほかの種の鳥の巣に産み、育てることもその親鳥に任せてしまうのだ。これについては後に詳述するが、育て親を騙すためにその種の卵とそっくりの色と形の卵を産むのである。カッコウはホオジロやオオヨシキリなどに、ホトトギスはウグイスに、ツツドリはムシクイ類に托卵するが、これら仮親の卵の色はチョコレート色、白に薄茶色の模様などまるで違う色をしている。カッコウ類はそれをちゃんと認識していてまちがいなく仮親が産むのとそっくりの卵を産みつける。驚くべきことは同じカッコウが託す仮親の卵でもホオジロ、オオヨシキリ、モズなどで違いがあるということだ。たださすがにカッコウの同じ個体が相手にあわせて産む卵の色を変えることはできず、ホオジロ型の卵を産むカッコウは一生その卵を産み続ける。その雛はホオジロに育てられ、自分が親になってもホオジロに托卵するから、カッコウには仮親に対応して遺伝的には別々の集団が存在するのである。さらに興味深いことに、ツツドリは本州ではムシクイ類に托卵し、その卵はムシクイ類の卵の白に近い白地に薄茶色の模様があるものだが、この同じツツドリがホトトギスのいない北海道ではなんとチョコレート色の卵を産むのである。ホトトギスはウグイスに托卵するためにその卵とそっくりのチョコレート色の卵を産む。北海道ではそのいわば競争相手であるホトトギスがおらず、托卵相手のウグイスの巣が「空いて」いるので「ウグイス型」の卵を産むのである。 仮親は巣の卵をあたためるが、カッコウの雛は少し早く孵ってほかの卵を巣の外に落としてしまう。そして仮親の運ぶ餌を独占して親よりも大きくなってもねだり続ける。この驚くべき一連の行動は、仮親が自分の雛を失った上に無駄な餌運びをするという意味でまちがいなくマイナスであり贋作といる。 ハチに擬態する昆虫類ハチは刺す。刺されると激しい痛みをともなう。現代ではハチに刺される経験をもつ人の割合は減っているが、それでもスズメバチに刺されるとニュースになる。ニュースにならない程度の軽傷は数多くあるに違いない。ハチが要注意であることは現代よりも自然と深くかかわりながら暮らしていた人々にとってよく知られたことであったろうし、それは他の動物にとっても普通のことである。 さてハチはしばしば黄と黒の縞模様を持ち、よく目立つ。このパターンは道路工事や広告にもよく使われ、人間の目にとまるものである。その結果、ハチに刺された経験のある人・動物はそれ以降ハチを見ると警戒するようになる。このことを利用して姿をハチに似て捕食をまぬがれる昆虫がいる。 昆虫類のハチ擬態は、ハエ・アブ類、カミキリムシ類、ガ類などで観察されている。ガ類ではスカシバガ科やカノコガ科によるハチ擬態が知られている。以下にはこれらハチ擬態の例をとりあげる。 アブのハチ擬態針を持たないのにハチの姿をまねている昆虫というと、まず双翅目のアブ類があげられる。森や林のなかを歩いているとき、アカウシアブが近寄ってきて体のまわりを旋回することがある。アブ類のなかでも大型のアカウシアブは、その飛び方、羽音、体色、体形がスズメバチそっくりで、まとわりつかれると一瞬身構えてしまう。ほかのウシアブ類もアカウシアブほどでなくても黒と白の縞模様のクロスズメバチにそっくりである。一般の人の目につくものとしては、花に飛来するハナアブ類があげられる。普通に見られるハナアブやシマハナアブはミツバチそっくりで[写真4a 左列]、虫に関心のない多くの人はミツバチと思っている。 またムシヒキアブ科のアブもスズメバチあるいはマルハナバチ類にそっくりである。大型のベッコウハナアブはマルハナバチそっくりで、アザミの花上などで見られる。ヨコジマナガハナアブは腹部の黄白色の横縞がクロスズメバチなどに似ている。フトハチモドキバエは体形と体色がスズメバチに似ており、とくに頬の縦長の褐色斑がそっくりである。ハチモドキハナアブは名のとおりハチそっくりで、ヒメバチ類に似ており、触覚の形までハチに似ている[写真4a 左列最上]。メバエ科には、触覚がさらにハチそっくりなのがいる。
カミキリムシのハチ擬態以下、上田・有田(1999)を引用しながら紹介する。「ハチをまねているのは双翅目だけではない。鞘翅目(甲虫目)では、ムネアカセンチコガネがマルハナバチ類の飛翔音に似た羽音を発するという。だが、鞘翅目でもっともハチ擬態を発達させているのは、カミキリムシ類であろう。たとえばトラフカミキリ類[写真4b 右から2列]やハナカミキリ類には、スズメバチやアシナガバチに巧妙に擬態しているものが数多くいる。ホソコバネカミキリなどはコンボウアメバチなどのアメバチ類に擬態している。ヒメコバネカミキリ類には、ヒメハナバチ類の一つの種にそっくりなものもいる。
カミキリムシは鞘翅目だから、前翅が堅い翅鞘に変化し、後翅は翅鞘の下に折りたたまれていて、飛ぶときにはそれが広がる。黒と黄色のハチ模様は翅鞘に出ている。それくらいなら、よく見ればホンモノのハチでないことが簡単にわかりそうだが、実はこれらのカミキリムシ類のハチ擬態はもっと巧妙である。かれらは後で述べるスカシバガ類と同様、その飛び方までがハチに似ている。とくにホソコバネカミキリ族のカミキリムシは飛び方がハチによく似ている。新大陸にはホソコバネカミキリ族の6つの特産属のカミキリムシが分布するが、北米の西海岸に生息するUlochaetes属のカミキリムシは驚くべきハチ擬態を見せる。U. leoninusのメスは飛ぶときに腹部先端を曲げてマルハナバチが飛んでいるように見せる。夏の終わり頃に出現するというのも、ハチの数が最も多くなった時期に合わせていると言える。この種は驚くと腹部を曲げて、しかも後翅を上にあげて羽ばたかせ、そしてハチのように刺す仕草までおこなうのである。」 スカシバガ類のハチ擬態(1)模様の擬態スカシバガ類のなかにはアシナガバチに似ているものや、スズメバチに似ているものなどさまざまなタイプが見られる。ヒメアトスカシバなどはドロバチ類に擬態しており、腹部に黄色や橙黄色の太い帯が1本から3本ある(上田・有田(1999))[写真4b 左下]。飛翔中はもちろんのこと、静止中もドロバチ類にそっくりである。一方、セスジスカシバなどは大型の種類でスズメバチ類に擬態しており、黒に幅広い黄色や橙黄色の縞模様がある。少し変わった例としては、カシワスカシバで、オスとメスとで色彩が異なる性的二型を示している(上田・有田(1999))。カシワスカシバのオスの腹部は黒に細い黄色の縞模様でクロスズメバチに擬態しており、メスは腹部はやや太い橙黄色の縞模様でアシナガバチや小型のスズメバチ類に擬態している。しかもこのカシワスカシバの成虫はスカシバガ類としては非常に遅く、9月下旬から10月上旬にかけて出現する。これはアシナガバチ類やスズメバチ類の個体数が最も多くなる時期であり、カシワスカシバは出現時期も「擬態」している。(上田・有田(1999)) 擬態とモデルについて興味深い事例がある(上田・有田(1999))。最近になって腹部にやや広い赤い帯のあるスカシバガが、北海道から一種(アカオビコスカシバ)と本州から別の一種(フクズミコスカシバ)相次いで発見された(有田、1997)。腹部の黒に赤い帯というのは非常に目立つ警告色であるが、モデルの赤い帯のあるハチは本州にはいるが、北海道からは見つかっていない。このためアカオビコスカシバはモデルが見つからないいわゆる「擬態やもめ」だという考え方もある。しかし擬態種の進化には、モデル種と擬態種が時間的・空間的に必ずしも同時に生息している必要はない。なぜなら北海道に生息する捕食者が本州やより南の越冬地でモデル種に会っていて、鳥がそのことを記憶していれば、こうした擬態の進化は起こりうるからである。 このように明らかにモデルが特定できるものもあるが、スカシバガ類全体としては、ハチ類の基本的パターンである黒と黄色の縞模様に擬態していると考えられている。というのは鳥にとっては黄色の帯の位置や数よりも黒の腹部に黄色の帯といった縞模様が警告信号として作用していると考えられるからである。 先にあげた「黒に赤という配色はドクチョウやドクガエルなど多くの毒をもつ動物に見られる普遍的な警告色パターンである。アカオビコスカシバやフクズミコスカシバが特定のハチの種をモデルにしなくても、この普遍的な警告色パターンに擬態していると考えればモデルのいない北海道に黒赤パターンの擬態種がいることの不思議さは説明がつく。スカシバガ類のハチ模様という形質は、スカシバガ類のほとんどの種において発現していることを考えると、これはもともとスカシバガの共通の祖先からの共有形質なのかもしれない。」(上田・有田(1999)) (2)体形の擬態このような一般的なハチへの擬態のほかに次のような特殊なものもある。オオモモブトスカシバやシタキモモブトスカシバはマルハナバチ類やミツバチ類に擬態しており、体全体が太くて重量感があり、後脚の脛節が長毛で膨らんでいる[写真4b 左上]。そしてその外側に黄色や橙黄色の長毛が密生して膨らんでいるために、まるでミツバチ類やマルハナバチ類が後脚脛節の花粉バスケットに黄色の花粉を集めて膨らんでいる状態によく似ている。日本で発見されたスカシバのなかにトックリバチ・ドロバチ類のような「腰がくびれた」ハチに擬態した種がいる。これらのハチは腹部基部(ハチでは腹柄と腹部第二節)が強くくびれて細くなっている。コシボソスカシバは腹部第一節と第二節がくびれたこれらのハチにそっくりになっている。ガ類では基本的に腹部第一節と第二節が太いが、コシボソスカシバでは細く変化しており、世界的に見てもあまり多くない。このような体全体の形態がイメージとしてハチに擬態しているのは、縞模様などの色彩が擬態しているのとはまた異なる標識的擬態と思われる(上田・有田(1999))。 (3)行動の擬態「スカシバガ類は形態だけでなく行動においてもハチとよく似たところがある。たとえばスカシバガ類は日中に活発に飛び回り、吸蜜のためにヤブカラシ、ハマセンダン、センダングサなどに訪花する。このときオオモモブトスカシバやシタキモモブトスカシバは花に飛来し、前脚を花にかけて空中でホバリングしながら吸蜜するが、そのときセイヨウミツバチやマルハナバチ類のように後脚を「く」の字に曲げる」(上田・有田(1999))ので、ハチのように見える。スカシバガ類の飛翔はどの種類も非常に力強くまた素早い。「羽音が捕食者に対してどのように伝わっているかははっきりしないが、少なくとも人間の耳にはハチ類とほとんど同じように聞こえる。 このようにスカシバガ類の多くの種類が、黒に赤や黄色といった縞模様の色彩だけでなく、ハナバチ類が花粉バスケットに花粉をためこんだような形態[写真4b 左上]や、「ハチのようなくびれた腰」という体形や、飛び方から羽音まで、総合的にハチ類に擬態している。」(上田・有田(1999)) シロオビアゲハによるベニモンアゲハへの擬態 ——進行中の進化を捉えるシロオビアゲハは東南アジア一帯から沖縄諸島にかけて分布するアゲハチョウ科の蝶の一種である。後翅の中央に白色斑文の列が見られることが特徴である[写真5]。一方,ベニモンアゲハも同じくアゲハチョウ科の一種で、マレー半島やインドネシア、フィリピンの諸島に分布し、後翅には赤色斑文が見られる[写真5]。両者は近縁種であり、色違いの同類のようだが、ベニモンアゲハは毒蝶であるのに対して、シロオビアゲハはそのような毒を持ち合せていない。この毒をもたないシロオビアゲハの擬態型のメスは毒をもつベニモンアゲハとジャコウアゲハ[写真5 右列下]をモデルとし、「シロオビ」アゲハとうその名に反して、後翅に赤色斑紋をもつものである。[写真5 右列中、下]注目されるのは、この擬態がメスに限られるということ、そしてわざわざ「擬態型の」という語を冠したように、シロオビアゲハのメスの皆がみなベニモンアゲハに擬態するというわけではないという点である。
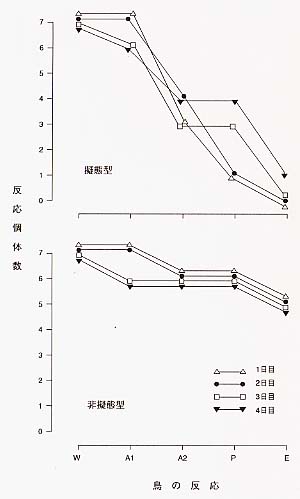
この擬態は捕食者である鳥など(信号受信者)にとって食用となるはずの種が、味がまずくて食用とならない種(モデル)をまねるものである。このような擬態はベイツ型擬態と呼ばれる。この型の擬態は、すべての個体が擬態するのではなくモデルの数に応じたある割合で擬態型と非擬態型とに分かれる。 さて、この擬態がなぜ雌だけに限られるのだろう。これについては大崎直太氏による優れた考察がある(大崎、1997)。ベイツ型擬態の多くは雌雄どちらも擬態型となるのが多いのに対して、蝶のベイツ型擬態はほとんどがメスのみに擬態型を発現する。この現象について大きく分けてふたつの仮説が唱えられてきた。 ひとつはグループ選択仮説である。あまりに擬態種の数が増え過ぎると、捕食者がこの蝶は食べられると認識してしまい、擬態種は食べられてしまう、だからすべての個体が擬態するのではなく卵を産むメスのみが擬態して、騙す効果を維持するのだという説である。もしこれが正しいとすると、オスは「メスのために」自らを犠牲にしていることとなる。しかし「利己的な遺伝子」(個体は自分の持っている遺伝子を最大限多く残せるように利己的に振舞うのであって、自分の遺伝子を犠牲にしてまで他個体の遺伝子を守ろうとはしないということ)の考え方が定説となったいま、この仮説は説得力を失っている。 もうひとつの仮説は、メスによるオスに対する性選択仮説である。オスは複数回の交尾が可能であり、オス同士のあいだでメスを巡って熾烈な競争がおこなわれ、自種らしきメスであればあまり厳しく吟味せずに交尾を試みるため、メスには変異個体も許容されるから擬態型も出現できる。これに対してメスは一度の交尾ですべての卵が受精可能なために一度の交尾が決定的で、相手のオスに対する吟味は非常に厳しいものとなるために、オスは祖先的な形態が維持されて、擬態型は出現しない、という考えである。つまりメスにとって相手のオスは正しいものでなくては困るが、オスにとってはメスは少々おかしくてもかまわない、だからメスのみに擬態型が存在し得たという説である。 このふたつの説を述べた上で大崎氏は調査に基づいて論を展開する。この調査では、毒蝶であるベニモンアゲハとシロオビアゲハのオス、擬態型のメスとが被襲撃率が低く、シロオビアゲハの非擬態型のメスは高い率にあるという結果が得られた[図1]。これは、擬態種にとってメスが擬態する効果は大いに存在するのに対して、オスはもともと捕食率自体低いので擬態する効果も小さいということを示している。しかしそれだけでは、少しであれ捕食率が下がるなら擬態型のオスがいないことの説明ができない。この疑問に対して大崎氏は擬態のコストと性選択説をとりあげる。
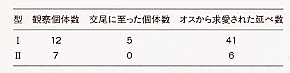
大崎氏による実験調査の結果、オスは非擬態型のメスを選ぶが、メスはオスを選ばないということがわかった[表1]。これは、性選択説の説く雌雄の関係とは正反対といってよい結果である。後翅にある白色斑文の列は通常型(非擬態型)のシロオビアゲハの特徴であり、擬態型のメスは赤色斑文をもつ分この白色斑文が減っている。ところがこの白色斑文こそがオスを引きつける要素であり、メスは赤色斑文をもっただけオスに選ばれなくなると考えられる。つまり擬態することは鳥に食べられないというプラス面がある一面で、オスに選ばれないというマイナス面もあることがわかったのである。さらにこの調査によれば、オスはどのような斑文型をもとうとも(さらに人為的に作った斑文でも)、メスに区別なく受け容れられたという。これはすなわち、オスには擬態することによるマイナス面こそあれ、プラス面はないということである。こうしたことをふまえて、このメスのみに見られる擬態においては、性選択説も受け容れることはできず、擬態することに対するコストという点からの把握が手がかりとなりそうであることがわかったのである。
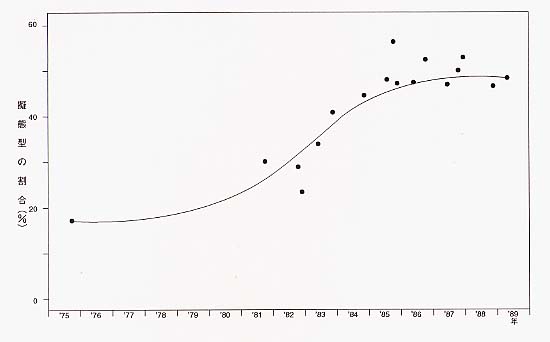
それでは、オスが擬態しないことで避けられているマイナス面とは何なのだろうか。これはそもそも雌雄にかかわらず擬態すること自体にあるものだと考えられるのだが、最近、この疑問を解く鍵が、沖縄諸島での研究からもたらされている。 もともとベニモンアゲハのいなかった沖縄諸島に近年ベニモンアゲハが分布するようになり、シロオビアゲハのメスの擬態型(赤メス)が多く観察されるようになった。この分布は地球温暖化が原因であると考えられるのだが、ベニモンアゲハが「迷蝶」、つまり、日本の外から季節風や台風に乗って飛んで来るというかたちで飛来したことに始まる。ベニモンアゲハは、1968年に八重山諸島に、1975年に宮古島に相次いで飛来、侵入し、その後定着することとなる。そしてベニモンアゲハの定着以後、シロオビアゲハの擬態型が見られるようになり、1975年以前には集団内で20パーセント以下だった擬態型(赤メス)が、1985年から1988年にかけてのあいだまでに50〜60パーセント増加した[図2]。以降、八重山諸島や宮古島では、擬態型の占める割合のほうが相対的に高い。これは、低率ながらも出現する擬態型が捕食者によって避けられ続けられた結果、本来の通常型よりも多くなったためと考えられる。
この沖縄諸島での事例を検証し、この擬態に関する重要な示唆をもたらす研究をおこなっているのが上杉兼司氏である。先述したように、擬態種はモデルの数に応じた割合で擬態型と非擬態型とに分かれるのだが、上杉氏が示した、ベニモンアゲハの多い島にはシロオビアゲハの擬態型が多く、ベニモンアゲハの少ない島にはシロオビアゲハの擬態型は少ないというデータは、このことをよく説明している。 さて、擬態すること自体のマイナス面だが、上杉氏が沖縄諸島でおこなった調査によると、毒蝶のいる島では擬態型・非擬態型の両型の寿命はほぼ同じだが、毒蝶のいない島では擬態型のほうが寿命が短いという。これは擬態型のほうが本来、生理的な寿命が短いということを意味しているだろう。このことは、擬態すること自体のコストとして生理的な寿命が短くなっているということを示唆するものだ。シロオビアゲハのオスは擬態する(赤オスになる)ことで避けられる危険よりも、このコストを払うことのほうがマイナスであるからこそ、擬態型が出現しないのだろう。 一方メスの擬態型は、オスに選ばれなくなることと、寿命が短くなることという、ふたつのコストを払っている。逆にいえば、擬態はこれほどの大きなコストを払って余りある利益をもたらすということである。 沖縄で見つかったこれらの事例は、この擬態が現在進行形で起こっているからこそ捉えられたものである。われわれは20年足らずで過半数近くに擬態型が増えたこの現象に対し、急速だという印象をもつ。しかし蝶の寿命は数ヵ月であり、百前後の世代を経れば、集団内の擬態型は相当数増加する。そのための20年というのは、十分な時間なのである。 このベニモンアゲハをモデルとした擬態は東南アジアの広い地域で確認されるのだが、シロオビアゲハの通常型は地域間の差が少ないのに対して、擬態型は地域差がはっきりと認められる。これは、擬態型が自分が分布する地域に居るベニモンアゲハをモデルとした擬態を見せているためである。ベニモンアゲハは各地域で少しずつ違った姿で分布しているので、それに擬態する擬態型もまた少しずつ違った姿を見せる。これらはみな、各地で何世代にもわたって擬態種とモデルの関係が続いた結果もたらされた現象である。従来、静的と思われていた種の組み合わせの分布が目の前で変化していくようすが時間を追って追跡されたという点で、沖縄諸島の事例は進化の動的な面を如実にものがたっている。私たちは擬態という現象に進化の妙を感じずにはいられない。 カッコウの托卵について托卵とは、ある鳥が他の鳥の巣に卵を産みこみ、その後の抱卵と雛の育成をその巣の主に任せてしまう行動のことである。そのなかでもカッコウ属(Cuculus)のカッコウ類は托卵という方法でしか繁殖せず、その托卵習性は最も高度に発達したもののひとつである。 以下カッコウ類の托卵習性について、(1)托卵相手の選択、(2)産卵行動、(3)卵の色と大きさ、(4)産卵数の4点について説明する。 (1)托卵相手の選択カッコウ類の托卵の相手は食物が同じでなければならない。これは雛の成長にかかわる問題で、食物が同じでなければ雛が順調に生育しないからである。次に、托卵する相手の個体数が多いという条件が必要である。なぜなら個体数が多ければ巣が見つけやすく、托卵の機会が大きくなるからである。また、相手の鳥が自分よりも体が小さいという条件が必要である。このことは托卵の機会を増やすことに関係する。概して小さい鳥は大きい鳥よりも高密度で生息していることが多く、托卵の機会が大きい。また小さい鳥に合わせて小さい卵を産めば、同じエネルギーでより多くの卵を生産できる。この条件は雛の生存とも関係している。カッコウ類の雛は、孵化した後で仮親の卵や雛を巣外に放り出す。それには相手の鳥が小さいほうが効率的である。実際日本で繁殖するカッコウ類4種、カッコウ、ホトトギス、ツツドリ、ジュウイチはすべて自分よりはるかに小さい鳥に托卵している。以上の3点が、カッコウ類の托卵相手に求められる条件である。ただしこれらの条件を満たす鳥であればどれでもよいというわけではなく、後に述べる卵の色や大きさなどにも影響されるため、通常は1種から3、4種を托卵相手として選ぶことになる。 カッコウ類の托卵相手は種によって異なる。これはカッコウ類の複数の種が同一の托卵相手を選べば、その托卵相手の鳥は過度の托卵を受けて個体数がいちじるしく減少し、カッコウ類の托卵の機会が減ることになるため、これを避けるためである。また、無秩序に托卵がおこなわれれば、一つの巣に複数のカッコウ類の卵が産みつけられるおそれが生ずる。一つの巣のなかで生き残れる雛は一羽だけであるから、他の雛が育つことができない。こうしたことから、カッコウ類の托卵は種間の競合を通しておこなわれてきたものと考えられている。 (2)産卵行動カッコウ類は托卵相手を巣づくりの段階から見つけている。でたらめに探すのではなく、樹上からじっと見張ったり、あるいは自分に対する相手の鳥の攻撃行動を逆に利用して、巣のありかの見当をつけている。そして相手が卵を産んだ直後に巣に行って卵を産みこむ。これは、もし抱卵期の巣に托卵すれば、雛が遅れて孵化し、生存の機会が減少するからである。カッコウ類のメスは産卵するとき、相手の卵を1個か2個くわえとる。産んだあとに抜きとってもよいようなものだが、カッコウ類の卵は仮親卵に外見が非常に似ていることが多いので、産んでからでは自分の卵を除いてしまう危険性があるからであろう。抜きとった卵は産卵中もくちばしにくわえたままでいることが多く、産卵後に食べる。この、卵を抜きとることの意義についてははっきりしない部分が多い。単純に数を合わせているだけとも思えるが、実験的に巣内の卵を1個や2個増やしてみても仮親は反応を示さないという。ひとつの可能性としては、仮親に自分を卵を食べにきた侵略者だと思わせて、托卵したことを悟らせないためであるという説明がある。もうひとつの可能性としては、現在のカッコウの祖先が巣内を独占せず、仮親の雛と一緒に育つ習性を持っていた時代のなごりがいまも残っているというものである。一つの巣の雛数は親鳥が育てうる最大の雛数になっている傾向があるので、自分の雛が一羽増える分、卵を一つ除くことは重要な意味があったのかもしれない。 (3)卵の色と大きさカッコウ類の卵は托卵する相手の卵に色や模様が似ていることが多い[写真6]。ただしカッコウにおいては変異に富んでおり、仮親卵に酷似しているものがある一方で、あまり似ていない例もかなりある。
カッコウ類の卵が仮親卵に似てきた理由の最も可能性のある説明としては、托卵された鳥が巣のなかに自分の卵と違う色の卵が入っているとそれを取り除いたり巣そのものを放棄するからであろうというものである。 このような、托卵される側の卵の識別能力も、托卵されてきた過程で托卵を拒否する手段として発達してきたらしい。たとえば、托卵されることのないカモメ類などでは、異卵を排除する行動は発達していない。カッコウ類が相手の卵にそっくりな卵色を発達させてきた背景には、托卵される側の識別能力の発達との熾烈な攻防があったに違いない。 カッコウ類のなかでも種や個体群によって仮親卵に対する卵色の類似に差が生ずる理由としては二つ考えられる。ひとつは先に述べたような托卵する相手の卵色に対する識別能力の差である。もうひとつの理由として、托卵するようになってきた期間の長さ、あるいはその隔離の程度が考えられる。托卵相手により長い期間にわたって托卵し、ほかのものからより隔離されてきたものほど卵の類似性を顕著に発達させてきたと考えられる。たとえば、本州中部を中心とした地域では、ここ20年から30年ほどのあいだにカッコウがオナガに托卵する例が増えてきた。カッコウの卵はオナガの卵とは似ていないが、托卵するようになってからの期間が短く、またオナガがカッコウの卵を除去することが少ないため、どの地域のカッコウもまだオナガに似た卵を産んだ例は確認されていない。これは托卵相手としての長い歴史をもつホオジロなどへの托卵が、見事な擬態卵を発達させているのと好対照である。(中村、1999) メスは同じ色模様の卵を産み、それは年が変わっても一定である。この類の雌雄関係が乱婚であることを考えると、卵色の決定には雌だけが関与している可能性が大きい。 一方、カッコウ類の卵は一般の鳥類と比較すると体の大きさに比べて小さくて軽い。このことの意義については次のように考えられている。卵が小さいと、抱卵の効率がよくなる可能性がある。仮親卵の大きさに合わせることにより、また卵を小さくすることで孵化に要する日数が短くなるだろう。さらに卵を小さくした分、また多数の卵を産むことができるだろう。ただ、托卵しないカッコウ類の特別に小さくはない卵でも孵化日数は同様に短いことを考えると、托卵するカッコウ類の卵が早く孵化することと卵が小さいこととは関係がないようである。 一方、カッコウ類の卵は仮親の卵よりはいくらか大きい傾向がある。これは、仮親に卵を受け入れてもらいやすくするためであると考えられる。一般に多くの鳥は、自分の卵より小さい卵を嫌う一方、大きい卵は積極的に受け入れる。たとえば、モズはスズメなど自分の卵より小さい卵は除去するが、自分の卵より大きいクロツグミの卵は受け入れたとか、ベニヒワはニワトリの卵やテニスボールまで温めようとしたなどの事例が知られている。カッコウはこうした大きな卵を好む鳥たちの習性をうまく利用するために大きな卵を産んでいるのであろう。ただ、あまり大きいと数多く産むことはむずかしいので、仮親卵より幾分大きめという現在の状態に落ちついたと考えられている。 (4)産卵数先にも触れたように、カッコウ類が一つの巣に一個しか托卵しないのは、雛が巣を独占するためである。実際、同じ托卵する鳥でも、雛が巣を独占しないカンムリカッコウ類やオニカッコウ類ではしばしば一つの巣に二個以上托卵することが知られている。カッコウ類が一繁殖期に産む卵数は、他の多くの鳥に比べて多い。これは一腹卵数が多いのではなく、繁殖回数が多いためである。その理由のひとつとして、卵が小さいことが考えられるが、カンムリカッコウのように特別小さい卵を産まないものでも卵数は他の鳥より多いことから、この理由だけでは十分説明できない。最も可能性が大きいのは、托卵習性そのものであるといる。つまり、カッコウ類は巣づくりや育雛などから解放されているため、それらへ費やすべきエネルギーを産卵数の増加へと注ぐことができるのである。 化学擬態人間は視覚に強く依存した動物だから、擬態についても形態に関するものが注目されてきた。しかしまねることはなにも視覚に限ったことではない。生物は生存をかけてあらゆる感覚に訴えて敵を騙そうとする。最近の研究によると、私たち人間にとっては驚くべきことがわかってきた(山岡・秋野、1999)。 アリの体表面についている炭化水素は種ごとに特異であり、アリが触覚でさわると同じ種であるかどうかがわかるのである。山岡・秋野(1999)はこれを「合い言葉」という表現を使っている。調べてみると,日本産の約50種はすべてがその種に固有な炭化水素の組成をもっていたが、一種だけ例外があった。それはサムライアリというアリで、このアリは少し変わった性質をもっている。サムライアリはほかの種の蛹や幼虫をさらってきて自分たちの巣に運び、そこで奴隷として働かせるのである。奴隷となるのであるから弱いアリかというとそうではない。さらわれてくるのはクロヤマアリやハヤシクロヤマアリなどで、違う種と出会うとはげしく闘ったり、逃げたりする。そのアリたちがおとなしく奴隷として働くとはどういう訳なのだろうか。 実はサムライアリは自分の炭化水素をできるだけ作らず、相手の奴隷となるアリに接触するなどしてそれを身にまとい、仲間として受け入れてもらっていることが判ったのである。人間からみえばみかけは違っていても、アリにとっては「合い言葉」である体表炭化水素のほうが重要なのである。その意味では奴隷となるアリは「あき盲」といる。 次の例はハチ、アリ、アブラムシ3種の組み合わせである(山岡・秋野、1999)。ハチはエイコアブラハバチ、アリはカワラトビイロケアリ、アブラムシはナシマルアブラムシである。アリはアブラムシと共生するが、ハチはアブラムシに寄生しようとするという複雑な関係がある。かれらの行動を観察していると実は奇妙な行動が見られる。アリはアブラムシをガードするから、寄生卵を産もうとしてアブラムシに接近しようとするハチにとっては邪魔となる。ハチはアリに何度も追い払われるのだが、そのうちアリのすきをみつけて突然馬乗りになり、後ろ足でアリの腹をさすると、アリはうっとりして動かなくなる。するとハチはさかんに触覚でアリの頭部を触れ、触覚をなめ、それから自分の体をグルーミングする。驚いたことに、この行動を30分ほど繰り返ししたあと、ハチは突然アリからはなれたかと思うと、自分の翅を噛み切って落としてしまうのである。こうなるとハチは飛べないからアリに攻撃されても素早く逃げることはできない。これではまるで丸腰、自分から銃を捨てたようなものだ。その翅なしバチは腹部を胸部側に折り曲げた奇妙な姿勢でアブラムシに近付く。もちろんアリは触角でハチに触れるが、どうしたわけかこのときはまったく攻撃をせず、ハチも触角で触れ返す。すると、あろうことかアリは攻撃しないばかりか、吐きもどして餌を与えるのである。ハチはこうしてしばらくアリの世話になって暮らしたあと卵巣の発達をまってアブラムシに寄生卵を産みつけることに成功する。 この謎めいた行動も最新のガスクロマトグラフ分析技術によって解明された。分析によると、馬乗りする前と後のハチの体表炭化水素組成はまるで違っており、それまでは木の葉の表面のワックスと同じn-アルカンのみにおおわれているが、馬乗り後にはアリのそれとまったく同じものになっていた。ハチは馬乗りしたわずかのあいだにアリの頭部から分泌される炭化水素を自分の体表に移したに違いない。こうなるとアリにとってはもはやハチではなく、仲間のアリそのものになってしまうのだ。 化学擬態というのはあまり馴染みのない現象だが、それは私たち人間の嗅覚が鈍感であり、あまり重要でないからに違いない。しかし多くの哺乳類や昆虫にとって、匂いははなはだ重要なものであり、ここに紹介したアリの例はその究極的なものといえよう。しかしそれでもこのような胸ときめく研究成果も偶然の産物という側面がかなりある。私たちが知らない「匂いの世界」は数限りなくあり、その世界で生きている無数の小さな動物たちがいるのである。 擬態について考える以上、動物によるいくつかの擬態について紹介した。しかし擬態は動物の専売ではない。ラン(蘭)の仲間にはハチにそっくりの花がある。この花はハチのメスに擬態しており、オスはまちがえて交尾しようとする。これを利用して受粉をおこなうのである。その花の写真をみたことがあるが、進化の妙といってもこれほどのものはないという傑作で、あきれてしまうほどハチによく似ている。しかし考えてみれば花はそもそも花粉を運んでもらうために植物が編み出した「装置」であり、その鮮やかな色彩、甘い匂いで昆虫や鳥やコウモリなどを引きつける。その極端な例がさきのランの花で、それはオスバチを欺いているから擬態といわれるのであって、その究極的な機能は送粉にある。多くの花はそのために嘘をつかないまでも「広告」をしている。その意味では本来緑である葉が変化してあれだけの可能な限りの色や形や匂いを生じた花は、自然界におけるとほうもない広告集団といる。そのように花をみるならば、そしてそのような目で自然界をながめれば、動物や植物は自分たちが生き延びるためにありとあらゆる手段で広告をしたり、欺いたりしていることに気づく。擬態はそのなかでもとびきりの傑作といってよいだろう。 私たちは自分たちのもつ感覚を自分たち以外のものにも延長しがちである。山に対して堂々とした印象をもち、嵐に怒りを感じる。機械類は働いていると感じるし、自動車や飛行機のような乗り物には明らかに生命を感じる。自動車の前照灯は目にみえるし、飛行機の翼は鳥のそれと対比させる。無生物に対してさえそうであるから、生物に対してはなおさらである。哺乳類ではいうまでもなく、鳥類や両生類でも目が視覚器官であり、鼻が嗅覚器官、口が食べるための器官であることは、子供でもわかる。母親はやさしく、子供はあどけないと感じる。それはごく自然のことであり、大半は正しくもある。しかしそのことを無批判に信じると事実を見誤ることがある。 わかりやすい事例はたとえばブッポウソウである。この鳥は「ブッポー」というような鳴き声をするのでこの名があるが、これがたまたま「仏法僧」と似ていたために信心深い鳥だと信じられていたが、これが生物学的にまったく意味がないことは明白だ。白く小さな花を清楚だと感じ、深紅の大きな花を妖艶だと感じ、竹に潔さを感じ、大樹に畏敬を感じる。しかしそれは心理的に自然で文学的に心引かれることであったとしても、それらの植物のもつ生物学的意味とはまったく関係がない。このようなことは明白なようだが、実は私たちが思い違いしていることはいくらでもある。 素朴なところではナマケモノを怠慢な動物だと思っている人はごく普通にいる。ハトは平和の象徴とされているが、厳格な社会をもっており、順位にしたがって優位の個体は劣位の個体をつつき、死にいたらしめることがある。逆に欧米で残酷な動物の代表とされ悪魔のように憎まれていたオオカミは劣位の個体を殺すことはない。パンダはこのうえなくかわいい動物と思われているが、飼育係が噛み殺されたことがある。それでも、それは例外的なことだと思われ、あのパンダが人を殺すなどありえないと信じられている。名前に由来する誤解はたわいがないとしても、外見からくる印象でステレオタイプなイメージを作ってしまうのは日常的におこなわれていることだ。洋の東西を問わず、どれだけ多くの動物が人間のイメージの延長上の意匠として採用されたことか。家紋や会社のロゴマークをみればそのことがよくわかる。生物学の歴史はそのような人間の価値観の延長による誤解を解いてきた歴史といるかもしれない。 イヌを飼った経験のある人はイヌにとって嗅覚がきわめて大きい意味をもっていることを知っている。確か散歩をしているイヌは匂いを嗅ぐことに忙しい。嗅覚だけでなく聴覚も大変鋭い。少なくとも人間よりはこれらの感覚がはるかに重要な働きをしている。こうした経験から、違う動物には違う感覚世界があるだろうことは想像できる。だがコウモリが人には聞こえない超音波を「聞き」ながら飛翔しているとか、人間には同じように見えるモンシロチョウの雌雄が、モンシロチョウの目にはまったく違う色に見えていて、オスはまちがいなくメスを見つけるとか、アリの行列は地面に残された匂いをたどっていることなどは近代生物学の成果を待たなければわからなかったことである。 このような例はおびただしくあるが、大切なのは自分たちのもつ感覚がヒトという特殊な種のものであり、ほかの生物にはまるで違う世界があるということに気づくことであろう。そのことを理解しながらも、カッコウの托卵にみられるようなさまざまな擬態は、相手を欺くことであるがゆえに、人間の倫理観が投影されがちである。食べられないように擬態している動物には「がんばっているな」とほほえましく感じたり、托卵のことを知り、雛が里親の卵を排除するのを見ると「なんてずるいんだ」と憤りを感じたり、その雛を育てる親を愚かだと感じたりしがちである。だがこれらはブッポウソウが信心深くないのと同じく、ほほえましくもなく、ずるくもない。擬態も生物の進化の必然的結果であり、生物が生存率を高め、死亡率を下げ、繁殖率を上げるために発展してきた目のくらむようなさまざまな生きざまのひとつのオプションにすぎない。ただ、それが温度が高いと植物の生産力が大きくなるといった物理・化学的に説明される現象とはあまりに違う、高度で簡単には説明できないものであることは確かだが。 (お断り——文中にいわゆる「不適当な表現」がある。しかしそれは擬態という興味深い生物現象を説明するためにふさわしい表現であるためで、他意がないことは文脈から明らかである。ご理解とご寛容を願いたい) |







【文献】有田豊「スカシバガとハチ擬態」、『生物科学49』、1997年、13−16頁。上杉兼司「シロオビアゲハが語る昆虫のベイツ型擬態の進化」、『擬態、騙しあいの進化論(1)、昆虫の擬態』、築地書館、1999年、73−92頁。 上田恵介・有田豊「黄色と黒はハチ模様」『擬態 騙しあいの進化論(1)、昆虫の擬態』、築地書館、1999年、62−71頁。 大崎直太「メスにのみ擬態型が現れるチョウのベイツ式擬態」、『生物科学49』、1997年、3−9頁以下。 高桑正敏『日本産カミキリ大図鑑』、講談社、1984年、27頁。 田中梓『日本動物大百科9 昆虫II』、平凡社、1997年、148頁。 中村浩志「日本におけるカッコウの托卵状況と新しい宿生オナガの托卵開始」《Japanese Journal of Ornithology,39》1990年、1−18頁。 樋口広芳『赤い卵の謎』思索社、1985年。 樋口広芳「托卵習性に見る鳥類の繁殖適応」、《Journal of Reproduction and Development, 41》、1995年、127−133頁。 樋口広芳『飛べない鳥の謎——鳥の生態と進化をめぐる15章 』、平凡社、1996年。 山岡亮平・秋野順治「アリ類の体表炭化水素の生態的意義」、『環境昆虫学——行動・生理・化学生態』(日高敏隆・松本義明監修、本田計一・本田洋・田付貞洋編)、東京大学出版会、1999年、479−491頁。 |
| 前頁へ | 表紙に戻る | 次頁へ |