熱帯林の地表・地中で活躍する社会性昆虫に関する海外調査 |
|
松本 忠夫 東京大学大学院総合文化研究科 |
私が熱帯の昆虫の研究を開始したのは一九七二年だから、早いもので、もうあれから四半世紀になる。最初に調査した国はマレーシアだったが、これは当時、世界各国で行われていた国際生物学事業計画(IBP)の一環で大学院生だった私が派遣されたものである。テーマはマレー半島の熱帯雨林の中でのシロアリ類の生物生産に関してであり、約半年の調査を一九七四年までに三回ほど行った。 派遣される前のIBPでの予備調査によって、すでに熱帯の生物生産を調べるに当たってはシロアリ類がたいへん重要なものであるという意見が出ていた。しかし、この当時、熱帯のシロアリの研究をしているプロの日本人はいなかった。また世界的にもまだ具体的な数値はあまり取られていなかった。特に東南アジアの熱帯林ではデータは皆無であった。そこで、そのような研究に着手する人材を育成しようということで公募が行われ、大学院生だった私と京都大学の安部琢哉氏(現在、生態学研究センター教授)がマレーシアに派遣されたのだった。 当時チームリーダーであった大阪市立大学の吉良竜夫先生や小川房人先生は太っ腹の人であり、ほんの駆け出しの私のような大学院生を長期間現地に派遣し、現地調査をやりたいようにやらせて下さった。これは本当にありがたいことだった。あの頃の体験から得たものは今でも身に染み着いている。その後、私は大学に勤めを得てからは、長期の出張はあまりできなかったが、タイ、インドネシア、台湾、オーストラリア、パナマ、コスタリカ、ベネズエラ、パプアニューギニアなどの熱帯のいろいろな国へ通算三十回以上も出掛けて現地調査を行ってきた。そして、熱帯のシロアリからさらに家族性のゴキブリの生態研究へと範囲を広げた。 最近の私の海外調査は毎年二、三回であり、たいていは研究室の大学院生とともに出掛けている。大学院生によっては単独で年に何度も海外に出掛ける者もいる。私が初めて海外に出た二十五年前に比べれば諸物価に比較して航空運賃はずっと安くなった。また現地の生活事情もずっと良くなっている国が多いので、現在は海外調査ははるかにやりやすくなったといえる。もっとも、未だに政治事情の悪い国あるいはむしろ悪化した国もかなりあるわけだから、私たちは単にそれらを避けているというだけであり、世界全体としてやりやすくなったというのは早計なのかもしれない。また、東南アジアなどではすっかり開発が進行してしまい、人手の入っていない原生林に容易にはアクセスすることができなくなってしまったのは残念である。
読者の中にはシロアリやゴキブリのような「嫌な虫」などが、なぜ海外学術調査の対象になるのかと、いぶかるむきもあろう。そこで、熱帯においてこれらの昆虫がどんな位置にあるのかを若干説明し、海外学術調査の対象としていかに魅力的であるかを知っていただきたいと思う。 日本では普段の生活の中でシロアリはめったに見られる昆虫ではない。熱帯でも多くの種類は地中や倒木の中などに棲んでいて、それらの生活はごくじみであり、またほとんど外界に姿を現さないので、見過ごされやすい。しかし、木造家屋は防蟻処置をしておかなければたちまち倒壊してしまうし、時期によっては羽蟻が大量に押し寄せるなどシロアリは活発な面も見せる。種類数は少ないが、高さ一〇メートル近くの巨大な蟻塚を作る種類、何十万頭という大集団で採食行進をする種類、大規模にキノコを栽培する種類などなど、温帯に住んでいる我々日本の居住者にはなかなか想像もつかない派手なものもいる[挿図1-3]。これらを実地で見た場合はだれでもシロアリ類を容易ならざる昆虫だと感心するであろう。
今までの学術的な調査から、シロアリ類はアリ類とともに熱帯における動物の中で現存量(面積当たりの生物重量)が最も大きく、その影響力のたいへん大きな昆虫と結論できるのである。 シロアリ類はハチ・アリ類などとともに社会性昆虫と呼ばれている。その理由は常に大集団で生息していること。その集団には女王や王と呼ばれるごく少数の生殖虫とワーカーや兵隊と呼ばれている多数の非生殖者がいること。そして、集団全体が高度に統制とれた行動をとることなどである。先に述べた蟻塚作り、採食行進、菌類栽培などはその典型例である。 シロアリ類の生態系の中での位置は、光合成をする植物を生産者、生きた植物を食う動物や肉食動物を消費者というなら、おもに死んだ植物を摂食しているので分解者ということになる。生産力の大きな熱帯林では常に大量の落ち葉や落枝あるいは倒木のような植物の枯死体が発生している。それらは地表部に堆積するが、セルロース、ヘミセルロース、リグニンを主体とした物質のため、菌類やバクテリアなら利用し分解できるが、通常の動物では食物にはできない。ところが、シロアリ類はこのような難分解性の物質を主要な食物としているがゆえに熱帯林中で大繁栄しているのである。その秘密は、シロアリの種類によって相手が異なるが、いずれも原生動物、菌類、バクテリアなどの微小な生物とのなんらかの共生関係を持っていることにある。ここでは詳しく述べる余裕はないが、シロアリ類が微小な生物と持っているこの消化共生の関係は生物界における種間関係の高度な例としてたいへん興味深いものといえよう。
私は熱帯のシロアリの調査をしていて、やがてこのシロアリ類の高度な社会性の起源がいったいいかなることにあるのかの興味を持ち始めた。時はあたかも日本において社会生物学がブームになりはじめた一九八〇年代の初期である。そして、その研究にとって、シロアリと同様に腐朽材を摂食するゴキブリが大変重要であることに気がついた。 このことはすでにアメリカのクリーブランドらが、古く一九三四年に指摘していた。彼らはシロアリの社会性の起源を考察する上で、北米のアパラチア山脈に分布しているキゴキブリ(Cryptocercus puncturatus )の生活の研究が重要であると唱えたのである[Cleveland et al. 1934]。このゴキブリは次のような特徴を持っている。
このキゴキブリの生活については、アメリカのナレパ女史によって最近の十数年間に一連の詳しい研究が行われた[最新の総説はNalepa 1995]。それらによると、キゴキブリの家族は下記のようなものである。
キゴキブリの後腸内には下等シロアリと同様、多数の種類の原生動物が生息している。まだ、詳細な研究はないがキゴキブリの場合も下等シロアリと同様に原生動物は食物の消化において、特にセルロース分解において重要な働きをしていると考えられる。つまり、これらの原生動物はキゴキブリと消化共生上の関係にあると考えられている。
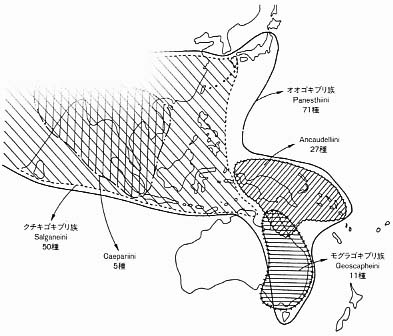
ここでいう食材性とは、森林の林床部にある倒木、落枝、木株などの枯死木(朽木)の中に生息していて、それらをおもに食物としていることである。世界で分類学的には現在までのところ、先に説明したキゴキブリ類(Cryptocercidae )と、オオゴキブリ類(Panesthiinae )の二系統がそのような食材性ゴキブリとして知られている。両類は外見上はいずれの種類もよく似ていて、成虫の体長は三−五センチで黒色であり、一般のゴキブリ類と比べると外骨格がたいへん堅く丈夫である。また、脚には刺を多数備えていて防衛に使われている。種類によって、長翅、短翅、無翅の違いがある。 キゴキブリ類とオオゴキブリ類は系統学的には離れていて、生態上の大きな違いは前者は卵生であり、後者は卵胎生であることである。そして、前者の方が原始的であると言われている。オオゴキブリ類は、世界的には東南アジアからニューギニア・オーストラリアにかけて百種類以上分布していて、典型的な熱帯性の昆虫である。日本においては、南西諸島を中心にして、オオゴキブリ属とクチキゴキブリ属(Salganea )の二属が分布している。前属にはオオゴキブリ(Panesthia angusutipennis )の一種、後属にはタイワンクチキゴキブリ(Salganea taiwanensis )とエサキクチキゴキブリ(S. esakii )の二種が知られている。近隣の台湾にはS. gressiti とS.raggei がいる[挿図4]。残念ながら、まだ大陸中国や朝鮮半島には他にどのような種類が分布しているのかの詳細な研究はない。
現在、私のところの博士課程の大学院生の前川清人君の手によって、本州、九州、南西諸島、台湾の順に弧状に連なった島々に生息しているこれらの五種類のゴキブリの系統関係が、ミトコンドリアDNAの分析により明らかになりつつある。それによると、種あるいは亜種レベルの系統関係と地理的分布とは見事に一致している。 前川清人君は日本列島ばかりでなく、東南アジアからオセアニアにかけての広範な種類の系統関係の解明を志している。
私はオオゴキブリ亜科の中でクチキゴキブリ属が、もう十年も昔になるが、家族性(亜社会性)の生活をしていることを見いだした。南西諸島および台湾で四種類を調べたが、どれも前節で述べたキゴキブリによく似た家族生活をしていたのである。子虫が成虫になるまでに数年間かかるが、その最終齢まで成虫とともに生活している。 面白いことに、同じオオゴキブリ亜科の種で日本列島に広く分布しているオオゴキブリ(Panesthia angusutipennis )の場合は、生まれた子虫はすぐ分散してしまって、親子間にはさしたる家族的な絆はうかがえない。 ところが同じ属の中でも、オーストラリアの亜熱帯雨林の朽ち木の中に生息しているスロアンオオゴキブリ(Panesthia sloanei )や、また温帯雨林にいる Panesthia australis を私が調べた場合には、それらの種では、子虫の若齢時には家族を形成していた。いったいどのような条件で家族が形成されるのか、また家族の持続期間はどのようにして決まっているのだろうか。次節で説明するが、おそらく卵から孵ったばかりの若齢期における体の発達の状態が、家族形成のあり方と強く関係していると考えている。 クチキゴキブリ類は卵胎生であり、産卵管から生み出された約二十個の卵は、直ちに雌の腹部の中にある卵嚢(保育嚢)の中に納められる。これらの受精卵はその中で孵化するまで大切に保護されているわけである。一般に親による子供の保護は、天敵や厳しい無機環境に対処する上で重要な行動である。このような卵胎生のゴキブリは、おそらくムカデやアリのような手強い捕食者から受精卵や若齢虫を護る意味があるのであろう。なお、生涯の産卵数が少ないことも、このような保護の様式の発達と大きく関連していると考えられる。 卵は卵嚢の中で数週間たったのち孵化し、一齢若虫となって外界へ生み出されていく。このような生まれたばかりの若齢虫は表皮は軟らかくたいへん弱々しい。タイワンクチキゴキブリでは、一齢幼虫は真っ白である。ところで、ゴキブリ類は不完全変態の昆虫であり、蛹の時期はない。そして、一般のゴキブリは初齢時からすでに脚が発達していて自力で歩行でき、また餌も取ることができる。このように早い時期から自立した生活ができることを早成性という。しかし、クチキゴキブリなどの場合は成長がたいへん遅く、なかなか自立できない。そのようなものを晩成性という。おそらく数年たってようやく成虫に達するものと思われる。ある程度自立できるまで、成虫は雌雄ともに子虫のそばにいる。このように両親がそろって子虫を保護する昆虫はあまりいない。食材性のゴキブリ以外には甲虫目のごく一部とシロアリ目で知られているにすぎない。 食材性ゴキブリの捕食者としては、朽ち木の中に潜り込んでいけるムカデ類が代表的なものである。特にクチキゴキブリやキゴキブリの若齢虫は、成虫による防衛がない場合にはたいへん弱い存在で、アリやクモなどにも容易に捕食されてしまう。オオゴキブリの場合は、さしたる親の保護行動がみられないが、それは、生まれてすぐの一齢幼虫でも黒色で小さいながら外骨格が丈夫であり、また大変すばしこく物陰にすぐ隠れることができ、捕食者から逃れるすべを持っている早成性であるからである。
成虫は種によって長翅を備えたものと、短翅のもの、そして全くの無翅のものがいる。長翅の種は成虫になってしばらくたった時に、交尾相手を求めて分散飛翔する。そして、雌雄が出会い、ペアー形成がうまくでき、そして朽ち木に営巣場所を得ると、両者は翅を失ってしまい、以後は飛翔できなくなる。タイワンクチキゴキブリの場合は、ペアー形成した雌雄が互いに相手の翅を食うことで翅を失う。この翅の食い合い行動は、果たして婚姻贈呈に相当するものなのか、あるいは相手の性を引き留めておくための手段なのか、たいへん興味深い行動である。エサキクチキゴキブリの場合は、最初から短翅であり、飛翔力は全くない。そこで交尾相手を求めるためには、もちろん歩行して分散せねばならない。食材性ゴキブリは長翅型の種類でも飛翔力は大きくないので、いずれにしてもそれらの行動圏はあまり広いものではないものと思われる。なお、オオゴキブリにおいては、翅は営巣木に擦れたりして破れて脱落するのみで、タイワンクチキゴキブリのように翅の食い合いはしない。 このような分散、求愛、交尾等は夜間に行われることであり、簡単に見ることはできない。雌雄および親子のコミュニケーションにおいて、たとえば誘因フェロモンなどを使用している可能性が高いが、まだ詳細は分かっていない。また、クチキゴキブリ類では実験室の飼育個体で、親が一齢虫に口移しに物質を与えているのを見つけたが、それがどのような物質であるのか、またどれくらいの量なのか、子虫にとってどんな意味を持つものなのかは、まだまったく分かっていない。
オーストラリアではサバンナ地帯の地中に成虫の体長が八センチもあるヨロイモグラゴキブリ(Macropanesthia rhinoceros )という極めて大型の種類が生息している。私はこれの子虫たちも約一年間成虫と共存し、家族生活をおくっていることを明らかにした。 このゴキブリの成虫は体長が八センチにも達し、生重は三〇グラムにもなる。[638][639]のように、上から見たところ、前胸背部が前方にまで発達し、頭部をおおっている。これはおそらく地中に潜る際に頭部を保護するとともに、スコップのような道具として使っているのであろう。体全体の外骨格は硬く頑丈で、六本の脚には鋭い刺が多数備わっている。不用意に手で握ったりすると刺がチクチクあたり大変痛い。人にたとえて言うなら、鎧を着て、兜をかぶっておまけに手足に武器を持った武者である。この武装はもっぱら防衛のためのものである。 前肢の先に備わっている刺はよく見ると、扇状に拡がっている。まるで哺乳類のモグラの手のようである[挿図5]。このような構造になっていることによって、土をかき分け、掘り、そして運ぶのに便利になっているのである。生息地の土壌は、砂質性であり、雨季は比較的軟らかくなる。ただし、べとつかない。その頃、この前肢とともに中肢と後肢そして前胸背をも使って、このゴキブリは地中深くにまで巧みに潜っていく。また、掘った土を体全体で押して地表部まで運び巣をつくる。あの防御に使われていると思われる鋭い刺は、実は穴掘りの道具でもあったのである。かれらは普段は地中四〇−五〇センチもの深いところにすんでいる。雨季になるといくら軟らかくなる土壌とは言え、この深さはこれらの道具なくしてとても掘り進めるところではないのである。
主な分布地はオーストラリアの東北部、すなわち雨季と乾季の明白な亜熱帯地帯であり、植生はサバンナや乾燥森林である。緯度は南緯一六度であるから、北半球であったならフィリッピンのマニラやタイのバンコックに位置する暑いところである。分布地が局限されているのは、おそらく先に述べたような土壌条件と強く関係しているのであろう。いくら丈夫な体をし力があったとしても、しょせん動物としては小さな昆虫であり、硬い岩石地や砂礫地では掘り進むことはできない。また、土壌の粒子が細かかったとしても、硬くしまっている場合には、やはりすみかにするには無理なのであろう。 私がこのゴキブリを研究したのは、少し前になるが一九八六年から一九八九年にかけてである。調査場所はクイーンズランド州の北部にある観光都市ケアーンズの近郊のマウントガーネットという優雅な名前の小さな町付近である[挿図6]。ここはオーストラリア全体の地図で見るとケアーンズ市のほんのすぐ近くに見える。とはいっても、日本の二十倍もある大きな国のことゆえ、実際に現地にたどり着くにはケアーンズ市から車でたっぷり半日はかかる。途中は標高が七〇〇−一二〇〇メートルのアサートン・テーブルランドであり、そこはオーストラリア有数の保養地である。そこから少し内陸側へ下ったところに、調査地は位置している。雨量の少ないこのあたりでは、川といっても水が流れるのは雨季の三カ月ぐらいのみで、普段はところどころに水が淀んでいるにすぎなく、乾燥の激しい年ではその水すらもなくなってしまう。あたり一面は、ユーカリを主体とした疎林であり、下草は主にイネ科草本である。乾季の終わり頃になると、緑の草はまったくなく、また野火で枯れ草は燃えてしまっている。このゴキブリは雨季・乾季の差の小さな日本から見れば、このような荒々しい風土の中に生きている。
このゴキブリは一生地中にいるわけではない。雨季にはかれらの食物である枯れ葉を取りに地表部に出てくる。ただし、日中には全く外に出てくることはなく、その採餌行動はもっぱら夜間に限られている。昼間に注意深く地面を観察すると、草の陰のあちこちに土が直径一五センチ、高さにして五センチぐらい盛り上がっているのが見える[挿図7]。そのようなところをスコップで掘ってみると、斜めの穴がポコッと出現する。この穴はたいがい弧を描きながら次第に深くなっていって一メートル以上は続き、長いものでは二メートルにも達する。そして、やがて深さにして四〇センチにもなると、土壌が急に硬くなり、巣穴は終点を迎える。その終点付近でゴキブリが発見される割合は七割ほどである。残りの三割はなにもいなかったり、大型のムカデが出てきたりする。巣穴の直径は平均八センチあり、その内面はスムーズである[挿図8]。このゴキブリはおそらく何度も土を運び出したり入ったりしながら、背中を穴の内面に押しつけスムーズにしているのであろう。そのような巣穴の途中にはたいがい枯れ葉が詰まっている。地表部に薄く堆積したユーカリ樹などの枯れ葉を引きずり込んで蓄えているのである。カラカラに乾燥した乾季は枯れ葉は枯れ草とともに燃えてしまって少ないから、この作業は雨季に行っているものと思われる。実際、このゴキブリが地表部を歩き回った形跡は雨季には見られるが、乾季には見られない。巣穴は場所にもよるが、一〇メートル四方あたり八十個も存在している。この高い密度から考えると、長い間には生態系の物質循環に相当な影響を与えているに違いない。
私はこのゴキブリの現地調査を三カ月おきに六回ほど行った。そして、全部で約三百の巣穴を掘り上げた。その結果、大変おもしろいことが分かった。雨季の始まる一〇月(現地の春)に巣穴の奥には、両親あるいは雌親とともに約二十匹の小さな一齢の子虫たちがいた。そして、これは次の調査時の一二月(夏)には三・四齢とすすみ、さらにその次の調査時の三月(秋)には六齢になっていた。ところがその次の調査時の六月(冬)には親虫と子虫たちが共存している巣穴はどこにもなかった。これらの調査結果から、このゴキブリは生まれてから六齢までの約半年は親と共存し、四月頃に親元から独立するようなのである。私はこの家族生活の様子を詳しく研究し、一九九二年に英語論文として発表した。 ところで、ヨロイモグラゴキブリも卵胎生である。また子虫と六カ月も共存している。これは親による子の保護が進化した例としてみることができる。そして、この場合、保護する理由は、乾季の厳しい気候条件と天敵の存在が考えられる。また、子供への給餌も重要な事項であろう。では、このゴキブリの天敵はどのようなものであろうか? 私は野外調査時に出くわした動物から、天敵はムカデ、大型のクモ、それにバンデイクートのような昆虫食の小型の有袋類であると考えている。大型のムカデが巣穴の中からしばしば発見され、実際、中齢の子虫にかぶりついているのを見たことがある。また、体長六センチもあるタランチュラのような大型のクモが付近の縦穴の中に生息しているが、ゴキブリの子虫を与えるとたちまちのうちに食べてしまった。ところで、親虫の方は外骨格はたいへん硬く、このような獰猛な捕食者でも歯が立たない。おそらく巣穴の途中に陣取って、外から来るムカデやクモのような天敵に対して体を盾にして子虫を守るのであろう。巣穴の中に食物貯蔵をする意味は、子虫たちが危険に身をさらしてまで採餌に出る必要がなくなることであろう。また、地中は温度と湿度が高いので菌類が繁殖し、食物の栄養価があがる可能性があり、これはある種の食物加工と見ることができよう。
今まで説明してきたような、ゴキブリとしては極めて特異的な形態と生活習性をもっているヨロイモグラゴキブリは、いったいいかなる祖先を持っていて、どのような理由であのように特殊化したのであろうか。その姿から、現地の人はこのゴキブリをたいへん原始的な昆虫と思いこんでいるようであるが、無翅、大型、卵胎生、家族性の発達などから考えると、むしろ、ゴキブリの中ではより進化したものと考えられる。 このゴキブリの場合は食材性ではなく、落葉食である。おそらくこのゴキブリは森林地帯に生息していた食材性の祖先から、サバンナ地帯へと進出し、その際、落葉食となり、さらに形態も特殊化したのであろう。食性とすみ場所や形態が変化しても弱々しい子虫がいるかぎり、親による子の保護行動はそのまま残ったのであろう。私はこのゴキブリの近い親戚は、食材性のオオゴキブリ属(Panesthia )であると考えている。このオオゴキブリ属は前に述べたように東南アジア、ニューギニア、オーストラリアの熱帯雨林に多数の種が生息し、主に腐朽した倒木や落枝を食物とし、かつそれを住処としている。また、その近縁の属に、やはり家族生活をしていて興味深いクチキゴキブリ属(Salganea )がいる。このようなオオゴキブリ属やクチキゴキブリ属とモグラゴキブリ属は系統学的にいったいどのような関係にあるのだろうか? それには形態や行動生態などに基づいた分岐分類学的研究とともにDNA分析が望まれており、私の研究室では現在これに挑戦している。 |







【参考文献】Cleveland, L. R., Hall, E. P., Sanders, E. P., Collier, J. 1934. The wood feeding roach Cryptocercus, its protozoa and the symbiosis between protozoa and roach. Mem. Amer. Acad. Arts Sci. 17: 185-342.松本忠夫、一九九三年、「シロアリの真社会性の起源とその維持機構」、松本忠夫・東正剛共編『社会性昆虫の進化生態学』、二六四‐二九七頁、海遊舎 松本忠夫、一九九五年、「シロアリの栄養生態学」『遺伝』四九、三一−三八頁 松本忠夫、一九九六年、「オーストラリアのサバンナの地中に家族で生活する巨大なゴキブリ」『昆虫と自然』三一、六−九頁 松本忠夫、一九九六年、「食材性ゴキブリの親子関係」『昆虫と自然』三一、二六−二九頁 松本忠夫、一九九六年、「シロアリの進化・ゴキブリとの関連」『木材保存』二二、二−九頁 Nalepa, C. A. 1995. Nourishment and the origin of termite eusociality. In: Nourishment and Evolution of Social Insects. (Hunt, J. H. & Nalepa, C. A. eds.) pp. 57-104. |
|









| 前頁へ | 表紙に戻る | 次頁へ |