




 |  |
| -20Kbyte image- | -32Kbyte image- |
デスモスチルスwスペルス Desmostylus hesperus
束柱目デスモスチルス科
北海道枝幸郡歌登町上徳志別、徳志別川中流河床ことぶき橋下流左岸約160m
地質調査所北海道支所地質調査員山口昇一発見、犬塚則久ほか発掘
1977〜78年発見@
医学部解剖学教室
ここに展示してある第2の骨格は、1977年夏に北海道枝幸郡歌登町(うたのぼりちょう)上徳志別(かみとくしべつ)を流れる徳志別川の河床で見つかり、翌78年との2年にわたって発掘された。産出したのは中期中新世のタチカラウシナイ層で、約1300万年前のものである。この化石は産地名から歌登標本といわれ、東京大学解剖学教室で古生物学的研究が続けられている。気屯標本は巨大な団塊から発見され、頭の前半、下顎、首の骨、前位胸椎、左右の膝、右の前後の脚が失われていた。いっぽう、歌登標本には、左の前後の脚の先と指の骨が欠けているものの、ほぼ全身が関節状態で埋もれていた。このため、日本の脊椎動物化石では珍しく埋没姿勢がわかる。歌登標本は第一大臼歯を使っている幼獣なので、体長165センチと小柄であるが、第二大臼歯を使う気屯標本は成体である。
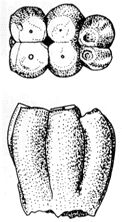 31-1 デスモスチルスの臼歯。上:咬合面、下:側面。
31-1 デスモスチルスの臼歯。上:咬合面、下:側面。
世界で最初にこの化石が見つかったのは1世紀以上も前のカリフォルニア州で、3咬柱分しかない臼歯の一部だったが、互いに密着しているので、デスモスチルスの特徴は表されている。そこで、この古代動物は歯を模式標本として1888年に命名された。デスモスチルスというのは臼歯の形に由来する属名で、ギリシャ語のデスモス(束ねたの意)とステュロス(円柱の意)を合成したものである。
分類学上の位置については、1900年代早々にデスモスチルスがゾウの仲間の長鼻類である、という別の意見が現れた。これは1898年に岐阜県の瑞浪市から世界最初のデスモスチルスの頭の骨が発見されたことによる。この頭骨の形が海牛類よりは、第3紀のかつて長顎マストドンといわれた長鼻類に似ていたからである。20年代になると、オーストリアの古生物科学の創始者、アーベルが多丘歯類や単孔類といったまた別の見解を表明した。このように異なる意見が続出する背景は、当初から、海牛類にも長鼻類にもおさまりきれない特殊性が、デスモスチルスに備わっていたからで、デスモスチルスを海牛目のなかにいれる人でさえ、その他の海牛類とは別の科や亜目という扱いをしていたほどである。 1933年には最初の全骨格である気屯標本が発見された。この骨格には頑丈な四肢の骨がふくまれていたので、海牛類とばかり思っていた世界の大多数の古生物学者は驚愕した。中にはその化石は別種のものが混ざったものであると考えた者さえいた。純水生の海牛類に脚があるはずもないからである。こうして、デスモスチルスは海牛類とは全く別の動物であることがはっきりしてきた。やがて、デスモスチルスの分類学上の位置は束柱目として独立することになった。昨今では長鼻目、海牛目、束柱目の3目はテチテリア類としてひとまとめにされる傾向にある。しかしそれでは、束柱目は長鼻目と海牛目のどちらにより近いのか、それとも長鼻目と海牛目が束柱目よりも互いに近縁なのだろうか。この3者のなかの親疎関係についてはまだ決着を見ていない。結局、デスモスチルスには分類学上は束柱目という名が与えられたものの、系統関係は1世紀以上にわたってまだ論争中ということになる。
 | 31-2 デスモスチルスの生体復元像(富山科学文化センター蔵)。肘や膝を張り出し、腹を引きずらんばかりに重心を下げている。 |
その理由は、化石の骨の形が奇妙であることに加えて、復元の方法論上の欠陥があったからである。つまり、それまでの復元法はといえば、まずその化石の分類をし、その種類に近縁の現生の動物の体形に似せて復元する、いわゆるモデル法であった。たとえば、日本からたくさん産出するナウマンゾウの復元の場合には、長鼻類には違いないので、現生のアジアゾウやアフリカゾウにならって復元して、そう大きな誤りは生じない。しかし、デスモスチルスのように、その分類上の位置そのものがはっきりしない動物では、モデル法でうまくいくはずがない。どの動物をモデルとして使うのかが決められないからである。
モデル法でゆきづまると、今度は制約のゆるい、いわゆる、関節法が使われた。この方法はもっぱら化石骨の関節面の形にしたがって、その形に忠実に組み立てるというものである。おかげで、モデル法による復元骨格に見られた関節での脱臼は姿を消したが、動物としてみたときに統一を欠いた不自然な姿が現われた。そこで、のちの人は誰も先人の復元姿勢を踏襲しようとはしていない。先人の作品に違和感を感じたからこそ、それをなくそうと、次々に別のスタイルを試みたのである。関節法では脱臼さえしなければ、各関節をどの角度でも設定できるので、様々な格好が可能である。こうして、たくさんのデスモスチルスの復元が生まれたが、同じ動物とは思えないほどバラエティーに富んでいたので、本当の姿はどうなのか、誰にもわからないまま最初の化石の発見から1世紀がたとうとしていた。
転機は、化石の復元に解剖学を応用することで、訪れた。この方法には骨の比較解剖学と骨と筋という運動器の機能形態学の2つがふくまれる。比較解剖学というと難しくきこえるが、要はほかの多くの動物との比較に基づいて復元することである。モデル法では特定のモデルにこだわったために、見解がわかれたが、関節法のように現生の動物をまったく参照しないわけにもいかない。そこで、デスモスチルスが所属することが明らかな哺乳類、とくに蹄をもった有蹄類をできるだけ多くモデルとして採用することにした。こうなると、それぞれの動物は体形がまちまちなのだから、ヌエや竜のように部分ごとに別の動物に類似させるのか、と思われるかも知れない。そうではなく、多様なモデル動物から共通点なり法則性なりを選びだし、哺乳類として、また、有蹄類としての共通基盤をもった、法則に則った体形に復元しようということである。
骨格復元の場合の法則性には、多くの動物にみられる共通の形質、骨と骨との間の角度、骨と骨格の関係、骨の形態と機能との相関の4種類がふくまれる。具体的には、脊柱の湾曲、首の長さと肩の高さとの調和、胸郭の断面形、体肢の方向、肢節の長さの比と運動機能、肢骨の関節の自由度、着地様式と中手・足骨、爪先の向き、前後肢の相似、があげられる。 比較の方法には、以上の法則性を抽出するための現生の哺乳類の骨格どうしの比較と、もう1つデスモスチルスの骨の特殊性をはっきりさせるための個々の骨の比較とがある。かねてよりデスモスチルスの骨は極めて形が変っているとされてきたが、各々の骨について、どの点がほかの動物とどう違うのかはっきりさせておく必要がある。さもないと、骨格復元が始まらない。骨格復元とは、体じゅうの骨にみられるその動物に特異な多くの点に、復元姿勢という単一の骨格形態から、解剖学的にみて合理的な説明を与えることである。つまり、化石の骨を単に骨格に組み立てたからといって、骨格復元をしたことにはならない。
さて、上にのべた哺乳類の骨格の法則性にすべての骨が従うとは限らない。どの動物にも、それぞれ特有の形態があるからである。そこで、2番目の機能形態学的な方法が必要となる。この方法は、体肢の骨の方向と体重を支える主な筋の働きとが調和するような復元姿勢を選択することである。体重支持筋は肩、胸、腰の周辺に付着するので、肢帯や基脚の骨に残る筋粗面や扁平骨の面積からその発達程度をうかがい知ることができる。
大きく分けて、4足動物の脚の向きは胴体の下か、横にはり出すかの2つである。両生類や爬虫類のように横にはり出す側方型では、体重が脚の開きを助長するようにかかるので、それに抗する体重支持筋は胴体の下から起こって大腿の内側や下面につく内転筋群が主体となる。反対に、普通の哺乳類のように脚を下に伸ばす下方型では、後肢の体重支持筋の主体は腸骨背面から起こって外転作用をもつ殿筋群である。
デスモスチルスでは前肢の上腕骨も後肢の大腿骨も外転位、つまり側方型に組んだほうが想定される筋の発達と調和がとれる。また、肘や膝を横にはり出すこの姿勢で組むと前腕の橈骨と尺骨の走り方、手根骨の形、膝の骨の大きさ、脛骨の内側へのねじれ、踵骨の先の傾き、足根骨の形などデスモスチルスに特異的な形象が合理的に説明される。この結果、側方型の姿勢が選択された。哺乳類の骨格構成上の法則性に対しては、体肢の方向だけが外れるが、その他のすべてには従っている。
こうして、デスモスチルスの体形は少なくとも上腕と大腿は胴体の下ではなく、横のほうにはり出していたことが明らかになった。先述の2方法で復元されたこの姿勢は、全身が関節状態で発見された歌登標本の埋没姿勢によって裏付けられることになった(挿図3)。歌登標本の化石は地層中で仰向けに倒れ、左右の四肢はそれぞれ胴体の左と右に分かれていたのである。もし、ウマやウシのように胴の下に四肢が伸びていたとすれば、死後海底に沈むときには左か右に横倒しになり、4本の脚は背骨に対して同じ側にそろうはずである。実際ほかの多くの哺乳類化石の関節状態の産状はそのようになっている。したがって、デスモスチルスの歌登標本の産状は、生きている時の体形を反映したものと考えざるをえないのである。
 | 31-3 デスモスチルスの歌登標本の産状図。仰向けに倒れ、左右の脚を開いている。 |
全骨格が発見されたことにより、完全な水生説は影をひそめた。しかし、両生説をとる場合には、主たる生活の場が水陸いずれにあったのかの検討が必要となる。たとえば、カバのように陸上で摂食して水中で休むのか、逆に鰭脚類のように、水中で摂食して陸上で休むのか、である。また、ひとくちに海岸といっても、三角州、マングローブ沼、潟、珊瑚礁などいろいろある。外洋か内湾か、砂浜か、磯かという点も解明しなければならない点である。ロコモーションや食性と合わせて考える必要がある。
もう1つの生息地に関する論争は北方系か南方系か、あるいは冷水性か暖水性か、という問題で、様々な見解がある。つまり、デスモスチルスは広範な温度差に耐えられたらしい、北太平洋地区の中新統に限られて産するので北方系の動物、共産する貝化石が冷水性なので寒流型ではないか、デスモスチルスやパレオパラドキシアがいた中新世は現在よりも温かかった、台島型植物群と密接に関連をもつので少なくとも最盛期は暖温帯の気候下に生息した、などである。
これらの根拠となる事実はいずれも正しい。逆の結論に達するのは、層準の精度、デスモスチルスとパレオパラドキシアという属の間の違い、海陸の温度差の問題もあるかもしれない。つまり、中期中新世の初めの短期間だけ温暖化した西黒沢海進期の影響とか、デスモスチルスが冷水性で、パレオパラドキシアが暖水性とする見方もある。デスモスチルスのほうがパレオパラドキシアよりも分布が北に偏っているのは確かであるが、ほとんどは時代も場所も重複している。両者が同じ露頭からみつかる場所もある。まだ解明を要する問題である。
水中遊泳の場合には、浮力を受けるうえ、前後の足の先が3次元空間を動くので、地上歩行ほど脚の動かし方に制約を受けない。いっぽう、地上歩行の場合には、立脚で体重を支えつつ、平面上を前進しなければならないので、脚の運び順や関節の屈曲角などの制約が水中よりは多い。そこで、まずデスモスチルスの歩行様式の復元をめざすことにした。
従来の研究では、デスモスチルスは骨格自体が特異な形なので、そのロコモーションもきっと変っていたに違いない、と考えられていた。しかし、ロコモーションの復元も現生の何らかのモデルを探すやり方ではうまくいくはずがない。骨格復元と同じで、ロコモーションのルールの探索と、その化石の特異性の抽出という、2つのアプローチが必要である。すなわち、まず多くの現生の大型哺乳類の歩容を調べ、歩容と体格、体重、骨の長さの比率など化石からでも分かるデータとの関連を見出す。化石のデータから、可能性のある歩容をいくつかに絞る。もう1つはデスモスチルスの骨格を主な関節が動くように組み立てて、想定される動きが歩行時に可能かどうかを確かめる。デスモスチルスは静止状態ではすでに側方型に復元されているので、いわば動的状態の復元をめざすわけである。
展示された歌登骨格は、このロコモーションの復元を目的とした関節可動式模型である。歌登標本には、脊柱をはじめ、前肢、後肢とも主要な骨が完全な形で残っているため、このような研究に資することが可能になった。この模型では、実際の動きを模して、体全体を左右にローリングでき、肩甲骨は胸郭上で左右に振ることができる。肩関節と股関節には3軸で動くユニバーサル・ジョイント、肘、膝、足首の関節は一軸性のヒンジ・ジョイント、手首には関節面の形から想定される複雑な車軸関節を備えている。動的復元とは、こうした哺乳類型の関節をもった動物が爬虫類のように肘や膝を横にはり出した姿勢で、実際にどう歩くか、という実験に当たる。実験の結果、前後肢とも背骨を蛇行させずに前後方向の軌跡を描けることがわかり、静的復元の確からしさが増した。いずれ動力を備えれば、歩く復元骨格も可能である。
食性の復元は古生態復元の中で最も難しい。根拠となる事実は多く、相反するものもふくまれているからである。形態学的に食性を復元するには、臼歯が最も重要で、次に切歯、下顎骨と頭蓋、全身の体形という順である。臼歯の柱状の咬頭と異常に厚いエナメル質は、圧砕に抗する構造であるが、磨耗に対しても強く抵抗する。いまでは、これらの仲間が海辺の砂浜にすんでいたと考えられているので、何を食べたにせよ、食べ物に混ざってくる砂がヤスリの役目を果たして、どんどん臼歯はすり減るはずである、と考えられている。こうした傾向に対する抵抗があの特異な臼歯のエナメル質を産み出したのだろう。祖先型のベヘモトプスから、パレオパラドキシア、コルンワリウス、デスモスチルスのどの進化型への臼歯の形態変化も強い咬耗に対する適応として説明できる。
下顎骨の形態も食性を反映している。歌登標本の発見で、はじめてデスモスチルスの下顎骨の完全な形態が明らかになった。この形をパレオパラドキシアや祖先型のベヘモトプスと比べることにより、食性の変化を探索できる。祖先型から進化型へは、下顎体・歯隙・下顎結合の伸長、歯列の短縮・後退という方向に進化した。特殊化の少ないパレオパラドキシアに比べるとデスモスチルスはさらに、下顎枝の前後短縮、関節突起の低下、下顎頭の後傾、下顎前端の上昇、臼歯の前傾という傾向が加わる。これらの特徴を現生哺乳類と比べると、束柱類はいくらか有蹄類に近い傾向を示し、デスモスチルスはパレオパラドキシアに比べて側頭筋よりも咬筋依存度が高いことがわかる。このことは頭蓋の矢状稜の有無や頬骨弓の太さによっても裏付けられる。したがって、基本的に束柱類は草食動物であり、デスモスチルスはパレオパラドキシアよりさらにその適応度が高い草食性である、と推定される。
(犬塚則久)