Abstract. As an example of cytological patterns in the Sino-Japanese flora, the karyotypes, chromosomal rearrangements, and fertility of Lycoris, a genus endemic to the Sino-Japanese floristic region, is discussed. The origin of two sterile Korean taxa, L. flavescens and L. chejuensis, is clarified.
Key words. Lycoris, karyotypes, hybrid complex, chromosomal rearrangement, Korean taxa.
Amaryllidaceous plants are found throughout the world except on Antarctica, but most genera are restricted to Latin America and South Africa. Only five genera are native to the Sino-Japanese flora and the genus Lycoris is endemic to this floristic region. Lycoris consists of about 30 taxa and can be found mainly in the laurel forest zone. This vegetation zone ranges from the eastern Himalaya to Japan, skirting southern Korea. Some taxa, however, such as L. aurea are found in the subtropical forest zone in Guangxi, Guangdong, and Hong Kong.
The systematic position of Lycoris is uncertain. Traub and Moldenke (1949) considered that the subtribe Lycorinae, consisting of Lycoris and Ungemia, and Griffinia, a member of the subtribe Amaryllinae, branched off rather early from the ancestral stock during the early evolutionary development of the family. Hutchinson (1973) treated Lycoris as a member of his tribe Hippeastreae. Kurita (1987) suggested that, from a cytological point of view, it might be closely related to Nothoscordum of the Alliaceae.
Although the chromosomes are small in size, the chromosome numbers and karyotypes in the genus Lycoris are very complicated, as are the flower shapes and colors. We would like to discuss these features in this paper.
Cytological patterns
Cytological studies of Lycoris were initiated in the late 1920s. To date, chromosome numbers and karyo-type analyses have been determined for most species, with the exceptions being only L. guangxiensis and L. shaanxiensis. Three major chromosome types have been found in the genus. One type comprises large metacentric chromosomes (M type). Another consists of telocentric chromosomes (T type), and the third contains acrocentric chromosomes (A type). In some taxa, these major chromosome types change their arrangement into other chromosome types (Kurita 1987, 1989). For example, Robertsonian fusion of two A type chromosomes of L. radiata results in a metacentric (M) and a small metacentric chromosome (m). An inversion in a T type chromosome in L. chinensis produces a submetacentric chromosome, sm (Fig. 1).
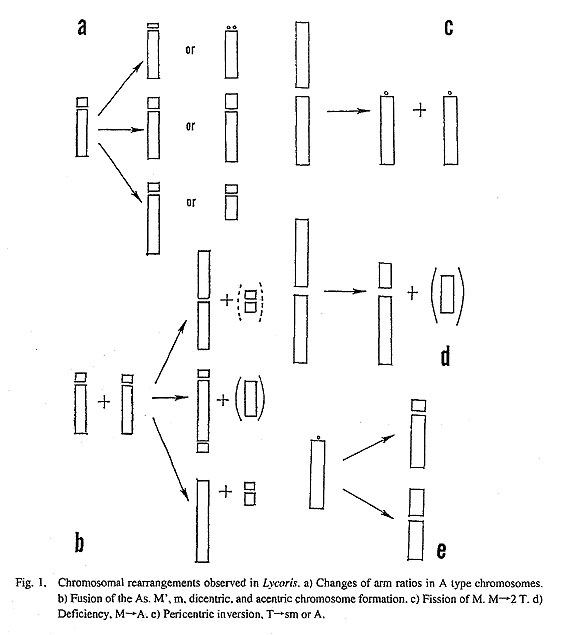
As shown in Table 1, the chromosome numbers and the karyotypes of the genus Lycoris vary considerably. The lowest number is 12, and consists of 10 M type and 2 T type chromosomes. The number of chromosomes increases gradually to 22. then jumps to 27, 29. 30, 33, and 44. The genomes of taxa having a relatively low chromosome number consist of M and T type chromosomes, while the genomes of taxa having 22, 33 and 44 chromosomes consist of A type chromosomes. Others consist of M, T and A type chromosomes. There are two kinds of genomes in this genus, however the fundamental number of both genomes is the same, 11. At a glance, this table presents many interesting facts, but in this paper we would
like to focus our attention on the relationship between karyotype and fertility.

Karyotypes and fertility
The number of fertile taxa, 16, is less than half of the total number of taxa for the genus. Table 2 shows the taxa known to be diploid and fertile. Taxa with an A type genome, 2n=22 A, are L. sprengeri endemic to China, L sanguinea and L sanguinea var. kiushiana endemic to Japan, and L. sanguinea var. koreana endemic to Korea. These taxa produce leaves in the early spring. Lycoris radiata var. pumila, L. rosea, and L. haywardii are endemic to China and produce leaves in the autumn. There are seven fertile taxa with an M+T genome, 2n=:6 M+10T, 8 M+6T, and 10M+2T. Lycoris chinensis is a continental species found in China and Korea. Lycoris anhuiensis, L. tongituba, and L. sperryi are endemic to China. Both L. traubii and L. aurea consist of 3 cytological races (cytoraces). The cytoraces of species with an even chromosome number are fertile. The former, with 2n= 12 and 14, may be found in Taiwan and southern Japan, and the latter, with 2 n = 14 and 16, occur in southern China.

More than half of the taxa of Lycoris are sterile (Table 3). There are no sterile diploid plants among species with an A type genome. Among the sterile diploids, taxa with an M+T type genome are L. traubii (2n = 13=9 M+4 T) and L. aurea (2n = 15=7 M+8 T). The remaining species have two kinds of genomes, M+T and A. Lycoris flavescens is endemic to Korea. Lycoris albiflora, L. rubroaurantiaca and L. elsiae are natives of Japan. The source of the four cultivated taxa is unknown, but they may be natural hybrids of Chinese origin. The rest may only be found in China. Among the sterile triploids, L. sanguinea is endemic to Japan, and L radiata is mainly found in China, although it may have spread to Korea, Japan and Nepal and become naturalized in prehistoric times. Among taxa with M+T+A chromosomes, L. chejuensis may be endemic to Cheju Island, Korea. Four others are Chinese taxa, among which L. squamigera is naturalized in Japan and Korea.

From a cytological point of view. most of the sterile taxa mentioned above may be interspecific hybrids. On the other hand, it was experimentally established that diploid species of Lycoris easily hybridize with each other, irrespective of their conspicuous morphological and ecological differences. Seed production in artificial hybrids is high and the seeds are easy to germinate and grow. Sterile F1 hybrids exhibiting meiotic failure can be maintained by means of active vegetative propagation of their bulbs, which probably explains why hybrids occur so frequently in nature. Many presumed hybrids are restricted to particular floras.
Fertile hybrids that propagate by seeds and bulbs result in polymorphic swarms of segregates and parental taxa, as are known in the L. sanguinea complex. It is not easy, however, to determine the actual parents of each natural hybrid taxon by cytology alone. Here, we would like to show two good examples of interspecific hybrid taxa found in Korea.
Parentage of the two sterile Korean taxa, L. flavescens and L. chejuensis
According to Lee (1980), 5 species of Lycoris, L. albiflora, L. aurea, L. koreana, L. radiata and L. squamigera, can be found in Korea. Recently, however, two more taxa, L. chinensis and L. flavescens have been recognized in Korea by Kim and Lee (1991). Moreover, Kim and Lee considered that the records of L. albiflora and L. aurea in Korea were attributable to misidentifications of L. flavescens and L. chinensis, respectively.
Karyotype analyses of Korean species of Lycoris were reported by Tae et al. (1987). Their results on L.
radiata, L. squamigera and L. koreana agreed with previous reports by many authors, but those on L. aibi-flora and L. aurea did not. We therefore reexamined the karyotypes of some Korean taxa collected by Dr. Muyeol Kim.
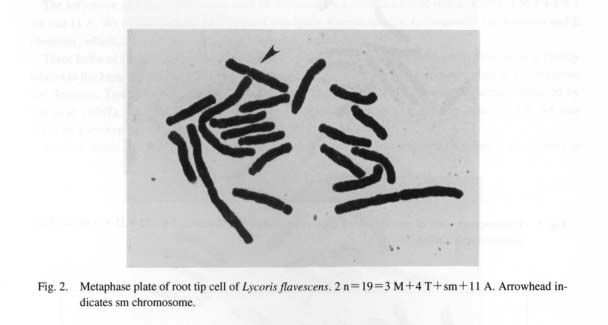
At first five bulbs of L. flavescens from Mt. Baekyang were examined. Lycoris flavescens is a sterile plant endemic to Korea and was once identified as L. albiflora despite the fact that its leaves sprout in early spring (Kim and Lee, 1991). Nineteen somatic chromosomes were observed. The karyotype consists of 3 large metacentric, 4 telocentric, 1 submetaeentric, and 11 acrocentric chromosomes (Figs. 2, 3). More precisely, the karyotype is formulated of 2n= 19=3 M+4 T+1 sm+11 A. The ratio of LM/SA, LM/ST, LA/SA, and r(sm) is 3.0, 2.6, 1.3, and 1.9, respectively (cf. Kurita, 1987). NOR was observed at the distal end of the short arm of sm. Such an unbalanced karyotype is usually thought to be produced by hybridization. This taxon is therefore not the same as plants studied by Tae et al. (1987) under the name of L. albi-flora, of which the karyotype was reported to be 2n = 18 and consisted of 4 metacentrics, 4 subtelocentrics and 10 acrocentrics. Tae et al. (1987) considered their plants to be diploids based on X=9.
The karyotype of Lycoris flavescens may be produced by a combination of two genomes, 3 M+4 T +1 sm and 11 A. We reexamined the karyotype of two fertile Korean species, L. sanguinea var. koreana and L. chinensis, which, from morphology, were assumed to be the parents.
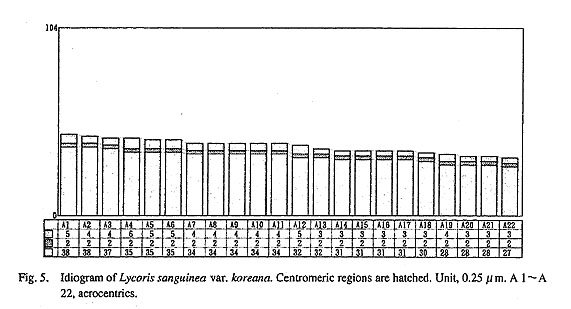
Three bulbs of Lycoris sanguinea var. koreana were collected from Mt. Baekyang. This taxon is closely related to the Japanese taxa of the L. sanguinea complex. Figure 4 shows a metaphase plate of L. sanguinea var. koreana. Twenty-two acrocentric chromosomes, 22 A, were clearly observed as already reported by Tae et al. (1987). One satellite chromosome was observed. As shown in Fig. 5, the values of LA/SA was 1.4. This karyotype agrees with the A type genome of L. flavescens.


2, submetacentrics.
Another fertile species, Lycoris chinensis was taken from Mt. Naejang. This species is also found in
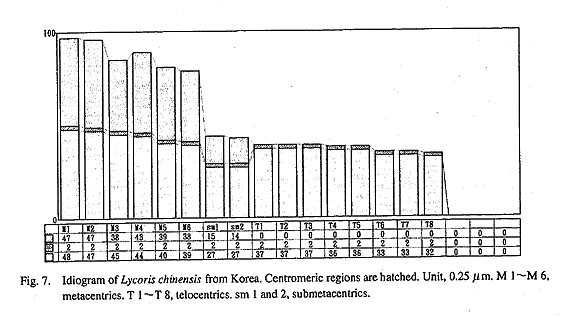
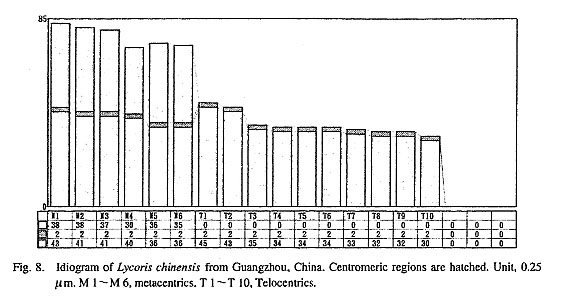
China. Figure 6 shows a metaphase plate of a root tip cell of the Korean L. chinensis. Six metacentrics, 2 submetacentrics, and 8 telocen trics, 6 M+2 sm+8 T, were clearly observed. The value of LM/ST was 2.9, and r(sm) was 1.9 (Fig. 7). Both values nearly agreed with those of the M+T+sm genome of L. flavescens. The submetacentric chromosome, sm, is very distinctive and is restricted to this Korean taxon. It has never been found in other species, not even in L. chinensis from China (Fig. 8), although Bose (1966) found a sm type chromosome in a bulb of L. chinensis of unknown origin. The sm chromosome might be produced by an inversion of the T 1 or T 2 chromosomes observed in the genome of the Chinese L. chinensis.

It therefore appears evident that the Korean Lycoris chinensis is one of the parents of L. flavescens. Namely, L. flavescens is a hybrid taxon between L. sanguined var. koreana and L. chinensis of Korea. Moreover, the karyotypes of the plants used in this study are nearly the same as those in plants examined by Tae et al. (1987) under the name L. aurea.
There is another sterile taxon which may be endemic to Cheju Island, Korea (Fig. 9). The shape of the flowers is very similar to that of Lycoris sanguinea var. koreana, although the flower size is larger. The brick-red tint of the tepals suggests a relationship with L. sanguinea var. koreana, while the somewhat crisped margins and pale yellow color of the tepal segments give hint at a relationship with L. chinensis.

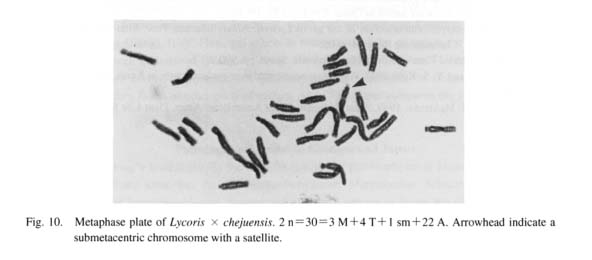
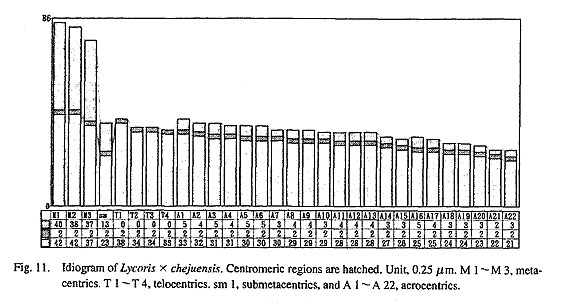
Figure 10 shows a metaphase plate of the plant from Cheju Island. The karyotype consists of 22 acrocen-trics, 3 metacentrics, 4 telocentrics and 1 submetacentric, 3 M+4T+ 1 sm+22A. The ratios of LM/SA, LM/ST, LA/SA, and r(sm) are 3.4, 2.5, 1.5, and 1.8 (Fig. 11), which strongly suggests that this plant is a trihybrid cross between L. sanguinea var. koreana, Korean L. chinensis and L flavescens from an unreduced gamete of the former fertilized by a reduced one from the latter.
This triploid sterile taxon is named and described here as a new hybrid:
Lycoris × chejuensis S.Kurita & P.S.Hsu. hybrid nov.
Foliis verno productis, 3-5, 30-40 cm longis, 7-1.5 cm latis. Umbella 5- vel 6-flora. Floribus tarde Julio usque ad Augusto productis, dilute aurantiacis laterici-vittatis, segmentis tepalorum 6-6.5 cm longis 1.2- 1.5 cm latis, tubo tepalorum 1.5 cm longo, staminibus quam segmentis tepalorum paulo longioribus; stylo perigonio paulo exsertiore. Scapus 50-60 cm longus, bracteis 2, lanceolatis, 3-3.5 cm longis, 1-1.5 cm latis; pedioello 1.5-4.0 cm longo. Bulbus ovatus, 4-5 cm longus, 3-4 cm latus.
Bulb ovate, with short neck, 3-4 cm in diam. Leaves similar to those of L. flavescens, appearing in February and persisting to May, 30-40 cm long, 0.7-1.5 cm wide at middle, apex rounded, tapering to 7-8 mm wide al base. Scapes late July-August, to 50-60 cm tall, light green, 8-10 mm in diam. at base. Umbel 5 or 6 flowered. Spathe valves 2, free, lanceolate, ca. 3.5 cm long. Perigone regular, margins of tepals slightly ruffled, particularly at mid section, orange-yellow, with a partial brick red band. Tepal tube 1.5 cm long. Tepal to 6.5 cm long, to 1.5 cm wide, linear-oblanceolate. Stamens slightly longer than tepals. Style overtopping stamens; stigma minute, nearly pointed. Karyotype, 2n =30=3 M+4 T+1 sm+22 A.
Type: Korea, Cheju-do, Mt. Halla, S. Kurita 81992 (flowers, holotype), 42093 (bulb and leaves, para-type) in Herbarium of Chiba University.
This paper presents only one example of the cytological patterns in plants of the Sino-Japanese Floristic Region. Cytological studies of eastern Asiatic plants have advanced rapidly in recent years, and chromosome number counts and karyotype analyses have been intensively investigated. Much more information is
still needed, however, for a fuller understanding of cytological patterns within this distinct floristic region. This paper is therefore intended to stimulate additional cytological studies of other groups of plants so that we may have a better understanding of speciadon and phytogeographical relationships within the Sino-Japanese Region.
Acknowledgements. We are indebted to Dr. Muyeo] Kirn of Chonbuck National University for his help in supplying bulbs of Lycoris from Korea.
References
- Bose, S. 1966.
- Karyotype alteration in L. chinensis. Sci. Cult. 32: 144-145.
- Hutchinson, J. 1973.
- The families of flowering plants, arranged according to a new system based on their probable phytogeny. Oxford Clarendon Press.
- Kim, M. Y. and Lee, S. T. 1991.
- A taxonomical study of the Korean Lycoris (Amaryllidaceae). Kor. J. PI. Tax. 21: 123-138.
- Kurita, S. 1987.
- Variation and evolution in the karyotype of Lycoris, Amaryllidaceae. II. Karyotype analysis of ten taxa among which seven are native in China. Cytologia 52: 19-40.
- Kurita, S. 1989.
- Karyotypic orftioselection in the genus Lycoris, Amaryllidaceae. Proc. Sino-Jap. Symposium Pl. Chromosome. PI. Chromosome Res.: 101-106.
- Lee, T. B. 1980.
- Illustrated Flora of Korea. Hyangmoonsa, Seoul, pp. 990.
- Tae, K. H., S. C. Ko and Y. S. Kim. 1987.
- A cytotaxonomic study on genus Lycoris in Korea. Kor. J. Pl. Tax. 17: 135-145.
- Traub, H. P. and H. N. Moldenke. 1949.
- Amaryllidaceae: Tribe Amarylleae. Amer. Plant Life Soc. California. 158-185.