Himalayan Potentilla and its Relation to the Sino-Japanese Floristic Region
Hiroshi Ikeda1,3and Hideaki Ohba2
1Museum of Nature and Human Activities, Hyogo 669-13, Japan
2University Museum, University of Tokyo, Tokyo 113, Japan
|
Abstract. Distribution patterns of common or allied species of Potentilla between the Himalaya and eastern Asia were surveyed. Four common species are shared between the Himalaya and Japan: P.fruticosa, P. anserina, P. nivea and P. sundaica. Their distribution patterns are discussed. Section Leptostylae, the most diversified group in Potentilla in the Himalaya, P. leuconota and P. tugitakensis are distributed in the alpine zone of Taiwan. Potentilla leuconota ranges from northwest India to southwest China (Yunnan, Sichuan and Guizhou) through the Himalaya and occurs disjunctly on Taiwan. Potentilla tugitakensis is endemic to Taiwan, and is similar to the Himalayan P. peduncularis. Although Taiwan is usually not included within the Sino-Japanese Floristic region, a considerable number of species from the Himalayan region, or with closely related species in the Himalaya, occur there. Key words. Potentilla, taxonomy, distribution. Himalaya. This paper discusses the distribution patterns of common or allied species of Potentilla between the Himalaya and eastern Asia. Potentilla is one of the most diversified genera in the Himalaya (Ohba, 1988). Table 1 shows the number of species distributed in Nepal, Bhutan, Tibet, the Hengduan mountains (southwest China) and Japan and also the four species in common between the Himalaya and Japan. These four species are: P. fruticosa L. in section Rhopalostylae, P. anserina L. s. lat. in section Leptostylae, P. nivea L. s. lat. in section Conostylae and P. sundaica (Bl.) Kuntze in section Potentilla.
Hara (1966) stated that P. fruticosa and P. kleiniana Wight et Arn. (= P. sundaica) are Japano-Himalayan elements known from the eastern Himalaya and Japan. Potentilla sundaica is distributed in the Himalaya, China, Korea, Japan, Sri Lanka, Sumatra and Java (Fig. 1). The habitat of this species, which grows in open moist places, such as on the banks of paddy fields and rivers, in the warm temperate region, is peculiar, because other species of Potentilla are found in cool temperate to alpine regions in the Himalaya. According to Hara (1966), most Sino-Japanese elements grow in moist places with mild climates. The pattern of P. sundaica is comparable with those of other Sino-Japanese elements. Although the distribution pattern of P. sundaica of the Himalaya, China and Japan is similar to that of typical Sino-Japanese floristic region elements, it is also distributed in SE Asia, which is outside the Sino-Japanese floristic region.
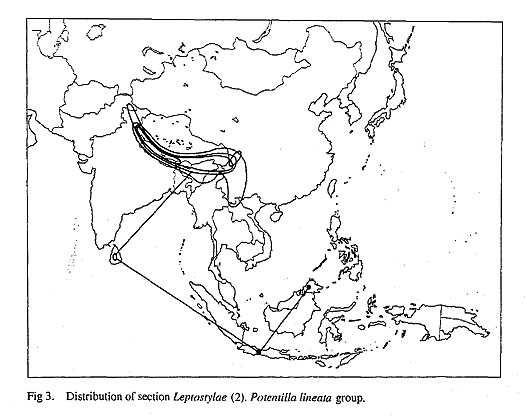
Potentilla fruticosa and P. nivea are similar in their distribution patterns. They occur in the arctic regions of Europe, the Alps, Siberia, the Caucasus, Mongolia, the Himalaya, Tibet, north and northeast China, Korea, Japan, Alaska, Labrador and western N America (Wolf 1908). The pattern of P. fruticosa and P. nivea might be interpreted as having spread widely throughout the northern hemisphere during the last glacial period and became reduced in range during the post glacial period. Potentilla anserina has a wider distribution than P. fruticosa and P. nivea, and is distributed not only from the temperate to arctic and alpine regions of the northern hemisphere, but also occurs in Chile, New Zealand, southeast Australia and Tasmania in the southern hemisphere (Wolf, 1908; Rousi, 1965). The distribution pattern of P. anserina requires a different interpretation than that of P. fruticosa and P. nivea. Rousi (1965) stated that the disjunct distribution of plants between North and South America could be ascribed to sporadic long distance dispersal (Raven, 1963) or to an ancient connection of the temperate floras between North and South America, which was made possible by a more continuous chain of high land masses than exist today in the Cordilleras and Andes (Du Rietz, 1940; Constance, 1963). She also speculated that the population of P. anserina on New Zealand (which she distinguished as P. anserinoides Raoul) reached New Zealand by means of long dis tance dispersal. The absence of indigenous representatives of the anserina group closer than Chile on the one hand or China and Japan on the other makes this a likely assumption. Since migration from Chile would not have involved crossing tropical areas, she thought migration from Chile to be more likely. The dispersal mechanisms and phylogeny of P. anserina and allied species need to be clarified before we can fully understand the origin of this pattern. Section Leptostylae Figures 2 and 3 show the distribution of section Leptostylae. The relationships of the species of section Leptostylae between the Himalaya and Southeast Asia is unknown. Potentilla leuconota D. Don and P. tugitakensis Masam. of section Leptostylae occur in the alpine region of Taiwan. Although Taiwan is not usually considered part of the Sino-Japanese region, but instead is placed in the South East Asian floristic region (Good, 1953), it is known that a considerable number of Sino-Japanese elements are distributed in Taiwan (Kanai, 1966; Hara, 1966).
Potentilla leuconota is distributed in northwest India, Nepal, Sikkim, Bhutan, Tibet, China (Yunnan, Sichuan and Guizhou) and Taiwan (Fig. 4). Potentilla leuconota of Taiwan is sometimes regarded as a dis tinct variety, var. morrisonicola Hayata, or as a separate species, P. morrisonicola (Hayata) Hayata. Hayata (1908) distinguished the Taiwan plants from var. leuconota by the absence of whorled leaves at the base of the flowering stems. We examined more than 200 specimens of P. leuconota collected from throughout the distribution range and concluded that var. leuconota is not distinguishable from var. morrisonicola by that or other characteristics (Fig. 5).
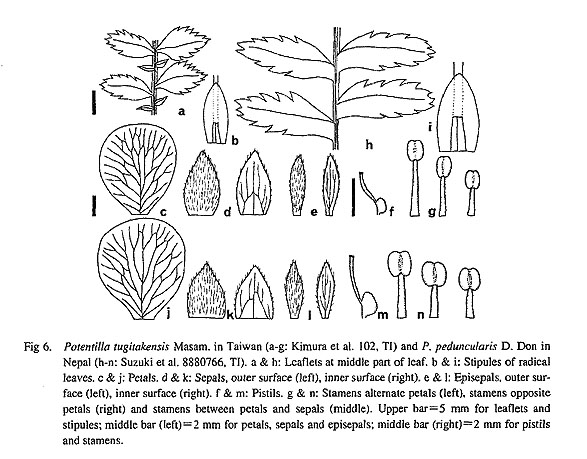
Potentilla tugitakensis is endemic to Taiwan. Although P. tugitakensis is sometimes treated as a variety of P. leuconota, var. tugitakensis (Masam.) Li, it apparently differs by the wholly connate stipule auricles of the radical leaves and the loose, non umbel-like dichasium as in P. leuconota. Ikeda and Ohba (in press) revised section Leptostylae in the Himalaya and adjacent regions and recognized five species groups within the section based on morphological characters (Table 2). Although it is not clear whether P. tugitakensis is related to the Himalayan species or to the Southeast Asian ones, e.g. P. borneensis (Stapf) Kalkman, P. tugitakensis is similar to P. peduncularis (Fig. 6) and may belong to the P. peduncularis group. Potentilla tugitakensis differs from P. peduncularis in the interrupted pinnate leaves. In Southeast Asia there are 17 species of Potentilla, of which 15 belong to section Leptostylae (Kalkman, 1993) and are found in alpine regions. It is possible that P. leuconota and P. tugitakensis or their ancestors migrated to Taiwan during the last glacial period.
To clarify the formation of the distribution of section Leptostylae, the whole range of distribution of the section in Southeast Asia needs to be examined. It is concluded that the species of Potentilla might expand their range by ways different from those of typical Sino-Japanese floristic elements. In regard to floristic region, it is interesting that the alpine flora of Taiwan contains a considerable number of species disjunct from the Himalaya. The formation of this distri bution pattern has not been thoroughly discussed. For P. leuconota and P. tugitakensis it can be assumed that they had a more extensive range across China during the last glacial period and now survive only at high elevations on Taiwan and in the Himalaya. Fig 6. Potentilla tugitakensis Masam. in Taiwan (a-g: Kimura et al. 102, TI) and P. peduncularis D. Don in Nepal (h-n: Suzuki et al. 8880766, TI). a & h: Leaflets at middle part of leaf. b & i: Stipules of radical leaves, c & j: Petals, d & k: Sepals, outer surface (left). inner surface (right). e & l: Episepals, outer surface (left), inner surface (right). f & m: Pistils, g & n: Stamens alternate petals (left), stamens opposite petals (right) and stamens between petals and sepals (middle). Upper bar = 5 mm for leaflets and stipules; middle bar (Ieft) = 2 mm for petals, sepals and episepals; middle bar (right) = 2 mm for pistils and stamens.
|






|
3 Present address: Faculty of Informatics, Okayama University of Science, Ridai-cho 1-1, Okayama-shi, 700-0005 Japan.[return to the text] |