Introduction
|
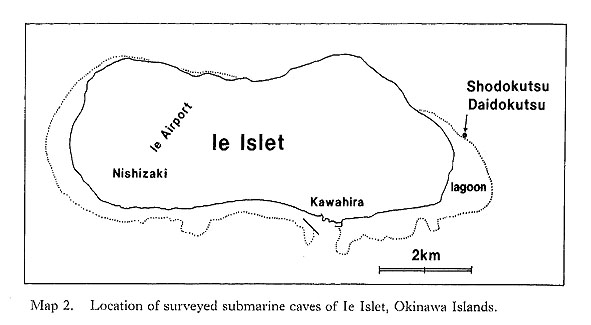
Submarine sheltered environments in and around coral reefs are often inhabited by characteristic and interesting organisms. Since Hartman and Goreau (1970) discovered sclerosponges (coralline sponges) resembling Paleozoic-Mesozoic stromatoporoids in the sublittoral crevices of Caribbean coral reefs, similar sclerosponge-bearing cryptic communities have also been found in the Indo-West Pacific and Red Sea reefs, and their composition, origin and evolutionary significance have been discussed by several authors (Jackson, Goreau and Hartman, 1971; Jackson and Winston, 1982; Maclntyre et al., 1982; Kobluk, 1988). Little, however, had been known about the feature of molluscs of such sheltered environments until we found living individuals of Neritopsis radula and a relict species of Pycnodonte from. the caves of the present area (Kase and Hayami, 1992; Hayami and Kase, 1992). The interior of lightless submarine limestone caves is one of the most interesting places for the study of cryptic organisms, but they were not accessible until recent years, when the technical development and popularization of SCUBA diving enabled scientists to explore for the first time the biota of this unknown world. Recently, with the assistance of several skilled divers, many interesting benthic organisms have been found in a number of submarine caves at Ie Islet of Okinawa Islands and Shimoji and Irabu Islets of Miyako Islands, southern Japan (Maps 1-3). They contain numerous undescribed bivalves and gastropods in addition to some benthic foraminifers, soft sponges, sclerosponges, ahermatypic corals, bryozoans, articulate brachiopods, polychaetes, a chiton (Lepidopleurus), crustaceans (ostracods, amphipods and decapods) and echinoids, which are to be described hereafter by cooperative and other specialists.
While taxonomic investigation has to precede advanced studies, the study of cave biota will produce various kinds of contribution to evolutionary biology. Because sheltered places may offer refuge to archaic and anachronistic organisms, submarine cavernicolous biota has been emerging as an exciting subject of systematics and paleontology. In fact, the present cave molluscs contain several "living fossils" as well as a large number of unfamiliar species with archaic appearances. We may gain a glimpse of ancient biota in such a cryptic environment. Submarine caves are very local and semi-closed places, but intensive studies of the biota may provide some generalization about the causal relation between the peculiar environment and the adaptive strategy of marine organisms. Several years ago, one of us (T. K.) learned that some lightless caves open to the sublittoral fore-reef slope of Ie Islet are inhabited by many individuals of Neritopsis radula, a famous bradytelic "living fossil" gastropod. At the same time it was found that the calcareous muddy sediments at the same site contain numerous molluscan shells of a unique assemblage. The preliminary reports (Hayami and Kase, 1991; Kase and Hayami, 1992) were based on the results of our field observation and laboratory investigation during 1989-1991. In 1991-1993, our study of cavernicolous molluscs was extended to a number of similar submarine caves of Shimoji and Irabu Islets of Miyako Islands. Meanwhile, an enigmatic "living fossil" oyster, Pycnodonte taniguchii, was found alive in several caves of these islets, as described in a separate paper (Hayami and Kase, 1992). The aim of this paper is to describe dominant bivalves from these caves and to discuss their evolutionary significance. Our systematic study, however, often encountered some difficulties. At present, anatomical study of these bivalves is not easy for us owing to their diminutive size, Nowadays, SEM observation and photomicrography of diminutive shells are rather easy, but close comparison with hitherto named species of similar size is often difficult unless the original materials have been examined through a similar procedure. In Japan and adjacent regions, greatly diversified marine bivalves have been known (Habe, 1977a; Higo and Goto, 1993), but our knowledge about the systematics of microscopic species and the ontogenetic development of larger species remains insufficient. The endemism of these species to cryptic environments often cannot be recognized on a firm basis. In some species we cannot necessarily deny the possibility that the cave material represents merely the juvenile stages of larger-sized non-cavernicolous species. The taxonomic identification and classification adopted in this paper, therefore, are often still provisional and will be improved by further studies. On the other hand, there are many advantageous points in the cave mollusc materials. The sediments on the cave floors are almost wholly composed of calcareous mud and bioclasts, containing numerous molluscan shells and other organic remains. They are almost free of terrigenous clastic material, indicating the sedimentation must be relatively slow. Most bivalve species are represented by numerous specimens. In cavernicolous environments physical, chemical and biological destruction (or erosion) of molluscan shells seems to be generally slow, and various characteristics in the early developmental stages (especially the external features of larval shells) are directly observable on the umbonal area of living animals and empty shells. This condition is suitable for the inference of larval ecology and developmental strategy. As enumerated elsewhere (Kase and Hayami, 1992) and discussed in some detail in the present article, the cavernicolous bivalves reveal several common characteristic features, which seem to give a clue for testing some theories of evolutionary ecology and marine biogeography. |