CHAPTER 15
The Animal Bones from the 1974 Excavations at Douara Cave
Sebastian Payne
Trinity Collage, University of Cambridge
| ( 5 / 10 ) |
|
15.2.3.20 Vulpes sp. (Plate 15.4) Vulpes is the most common carnivore at Douara Cave; measurements are as follows:
Vulpes vulpes is very widely distributed in the Near East, both in the desert zone (though probably not in the center of the great sand deserts [Harrison, 1968]), and in the wetter areas to the north and on the Levant coast. Size variation is considerable, foxes in the desert zone being smaller than those to the north; the Douara Cave specimens are relatively small in these terms. Davis (1977) suggests a correlation between size and temperature, interpreting size reduction in Israel at the end of the last glacial as a direct effect of temperature increase; an alternative possibility, however, is that the size reduction reflects the replacement of a larger, northern ecotype by a smaller, southern, semi-arid ecotype. 15. 2. 3. 21 cf. Fennecus / small Vulpes (Plate 15. 4b). A very small fifth metacarpal is clearly canid rather than felid, mustelid or viverrid; it compares very closely with V. vulpes, but is much smaller. Measurements are as follows:
This is about the same size as Fennecus serda, judging from two specimens in the British Museum. (Natural History) collection; detailed comparison was not possible, as the specimens are still in articulation, with the cartilage not removed. In the absence of suitable comparative material of Fennecus and of Vulpes ruppelli, the identification of this specimen must remain uncertain. Both are small desert foxes, found in the Near East and in North Africa; V. rüppelli is fairly widely distributed in desert and semi-desert areas, while Fennecus is typically found in sandier habitats. Neither has been reported from cave sites in the Near East. Felidae Felid bones are relatively scarce in the Douara Cave collection: 11 specimens were found, and at least three species are represented, one very large, one of medium size, and one relatively small. A number of species occur today in the Near East or have occurred recently; these include: Panthera pardus, the leopard; P. leo, the lion; Acinonyx jubatus, the cheetah; Felis caracal, the caracal lynx; F. chaus, the jungle cat; F. lybica and F. silvestris, the wild cat group; and F. margarita, the sand cat. 15. 2. 3. 22 Panthera cf. leo (Plate 15. 5)
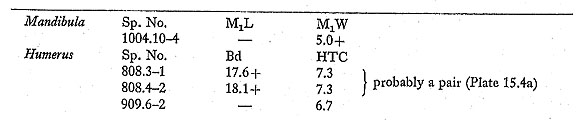
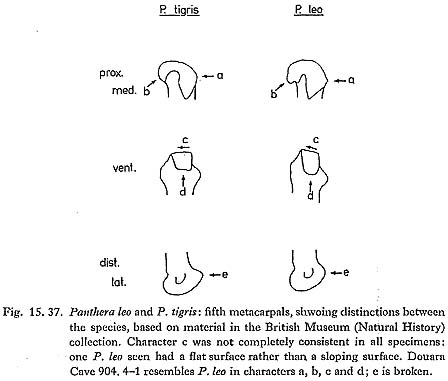
A fifth metacarpal is very large and robust. The characters shown in Fig. 15. 37 separate P. leo (lion) and P. tigris (tiger) on the skeletons in the British Museum (Natural History) collection, and the Douara Cave specimen is referred to P. leo on this basis. Two milk teeth and a burnt juvenile mandible fragment are again from a very large felid, and in all probability from the same species; an unfused phalanx 1 and an unfused radius, both from very young animals, are more tentatively referred to the same species. Measurements are as follows:
Lions are probably now extinct in the Near East; they survived into this century (Harrison, 1972: 621). Perhaps surprisingly there appear to be no records of lions from Pleistocene sites in the Near East except perhaps from Oumm Qatafa (Vaufrey, 1951), although there are reports from more recent sites, and the lion is of course mentioned in the Bible. 15. 2. 3. 23 Felis cf. caracal (Plate 15. 5) An innominate compares closely with the only specimen in the British Museum (Natural History) collection, especially in having a somewhat depressed rectus attachment area, and differs in this respect from other felids of similar size. The British Museum (Natural History) specimen (Reg. No. 1855. 9. 17. 2) is, however, badly deformed and cannot be assumed to be typical. A tarsal is from an animal of similar size; no comparatives were available. The only measurement is as follows:
The caracal lynx is rare but widely distributed from Africa to India; it is found in mountainous and hilly steppe country, and does not penetrate far into sand desert areas (Harrison, 1968). 15. 2. 3.24 Felis sp. (smaller) Three specimens are from smaller felids: two ulnae are about the size of a large F. silvestris, while a burnt edentate mandible fragment (905. 3-8) appears to be from a some-what smaller animal. Measurements are as follows (only one of the ulnae could be measured):
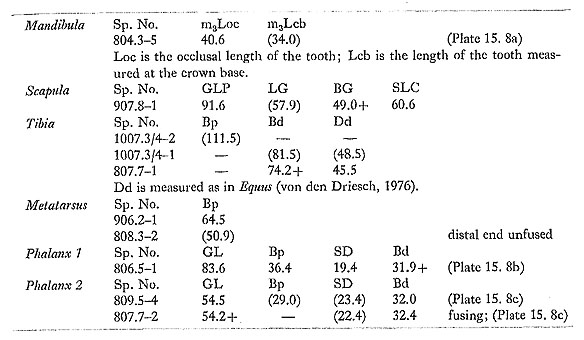
PERISSODACTYLA Equidae 15. 2. 3. 25 Equus hemionus / asinus (Fig. 15. 38; Plates 15. 6-7)
Teeth and tooth fragments of a small equid were fairly common; bones were somewhat scarcer. Crown patterns of the more complete teeth are illustrated in Plate 15. 6. Two permanent upper cheek teeth, in full but relatively early wear, both have caballine folds. The mesial and distal walls of the fossettes are moderately folded; in the one complete specimen the styles are relatively heavy and buccally flattened, and the interstylar walls are flatly concave. Measurements taken on the complete tooth give a protocone index of 44.5:
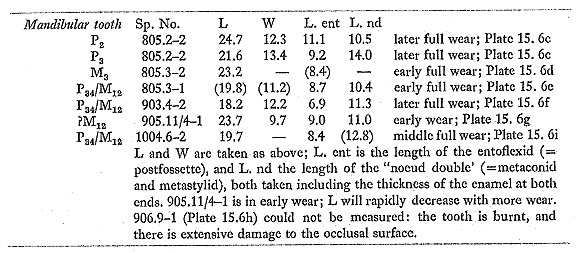
Eight lower permanent teeth include two mandible fragments (possibly from the same individual), one with P2 and P3, and the other with M3. Lingual sinuses are variable, from shallow and irregular (906. 9-1: Plate 15. 6h) to U-shaped (905. 11/4-1: Plate 15. 6g) or fairly sharply V-shaped (903. 4-2: Plate 15. 6f). This may to some extent be related to the state of wear: V-shaped sinuses tend to occur on teeth that are more heavily worn. Penetration of the buccal sinus is again variable, from shallow to moderately deep. None of the teeth has more than a trace of a ptychostylid; entoflexids are generally simple and asymmetric, with no more than a trace of a hypoconid fold. The following measurements were taken:
There are two milk molars. One is hardly in wear; the other is a lower milk molar in early full wear, with a well developed ptychostylid (Plate 15. 6j). The following postcranial measurements were taken:
At least three equid species can be identified from Upper Pleistocene sites in the Near East. Equus caballus is present but scarce at Mount Carmel (Bate, 1937), at Ksar'Akil (Hooijer, 1961), and perhaps at Palegawra (Turnbull and Reed, 1974: 116). E. hydruntinus is reported from a number of sites, and Davis (1980) regards the larger E. mauritanicus from earlier sites such as Djebel Qafzeh as possibly ancestral to E. hyduntinus from the later sites. Finally, an equid of the E. asinus / hemionus group has been reported from a number of sites, generally in drier areas (Davis, 1980). This equid has traditionally been identified with the recent E. hemionus hemippus of northern Syria, but Ducos (1970) has questioned this, pointing to the scarcity and unsatisfactory nature of the available material of E. h. hemippus and suggesting, on the basis of a large equid sample from the early Holocene site of Tell Mureybit (northern Syria), that wild E. asinus may have extended into the Levant. Eisenmann (1980) rightly stresses the considerable variation shown by equid cheekteeth, as well as the importance of working with large samples and with teeth whose position in the tooth row is known or can be established. The equid material from Douara Cave falls far short of such requirements. At the same time, there is nothing in the Douara Cave material to suggest that more than one species is present, and the characteristics of the material fit well with the E. asinus / hemionus group: this is suggested by the lower cheek-teeth (shape of lingual sinuses and relatively shallow penetration of the buccal sinus), and by the slenderness of the two complete metacarpals (SD/GL=11.8% and 12.3%; compare Bökönyi, 1972; Eisenmann, 1979; Davis, 1980; and Fig. 15. 38). Distinguishing between E. asinus and E. hemionus is more difficult, and Eisenmann (1979; 1980) stresses their close relationship. The size and proportions of the Douara Cave metapodia are close to those of recent Syrian onager (Fig. 15. 38), but Ducos (1970) has questioned what this was. A simple choice between E. hemionus and E. asinus is not the only possibility: Davis (1980) suggests that the two may possibly be a single species, changing clinally from Africa to Mongolia, and that "if this is correct, then an Israeli wild ass would have been morphologically intermediate between the Egyptian ass and the Iranian half-ass," A further possibility is that differentiation from a common E. asinus / hemionus stock may have progressed further than that, and that the small Levantine form may have been a separate species, just as Eisenmann (1979; 1980) suggests that E. tabeti, a larger species from Pleistocene sites in North Africa, may have been another offshoot from this common stock. ARTIODACTYLA Camelidae 15. 2. 3. 26 Camelus sp. (Plate 15. 8)
Camel bones were quite common; the following specimens were measured:
As the measurements show, these were not particularly large camels: they are comparable in size to modern domestic camels. Camel remains have been reported from a number of Upper Pleistocene sites in the Near East, but rarely in any number. There are fragments from Emireh (Bate, 1927), Sahba (Vaufrey, 1951), Qafzeh (Bouchud, 1974), Far'ah II (Gilead, 1980; C. Grigson, personal communication), and Tabun (Garrard and Payne, in preparation), and more abundant remains from C-Spring, Azraq (Clutton-Brock, 1970). The Sahba and Azraq camels were identified as C. dromedarius, the Arabian camel, but Bouchud is undoubtedly right in warning that caution is needed: there are as yet few reliable osteological criteria for distinguishing C. dromedarius from C. bactrianus, the Bactrian camel. While the Azraq and Douara Cave camels are of medium size, the Far'ah camel is very large (C. Grigson, personal communication); considerable variation can be expected within a population of camels, but the very large difference between the Far'ah camel and the others raises the possibility that there may have been more than one species of camel in the Near East during the Upper Pleistocene and that the large Far'ah camel may be related to C. thomasi, a large camel from the Pleistocene of North Africa, which Gautier (1966) suggests is closer to C. bactrianus than to C. dromedarius. Shortage of material makes these problems difficult to resolve; but what is undoubtedly needed at the outset is a detailed study of osteological differences between modern C. dromedarius and C. bactrianus. Bovidae 15. 2. 3. 27 Alcelaphini, gen. et sp. indet. (Plate 15. 8d) A lower molar (1009. 2-1; Plate 15. 8d) is clearly alcelaphine, on the basis of the rounded buccal lobes, complex infundibula, and absence of a basal pillar or a "goat fold" (Gentry, 1978); it has an occlusal length of 22,0 mm. Similar specimens have been reported from several Upper Pleistocene sites in the Near East (e.g. from Mount Carmel; Bate, 1937: Fig. 7g), and are usually referred to Alcelaphus, mainly on the basis of reported sightings near the Dead Sea during the last century (Tristram, 1882). But teeth of other alcelaphines such as Beatragus and Damaliscus are very similar, and it seems prudent not to identify the Douara Cave specimen more closely. Four other fragments from ungulates of about the same size compare reasonably with Alcelaphus and other alcelaphines, but cannot be definitely identified as alcelaphine: an upper molar fragment (805. 5-3), a scapula fragment (1008. 11-3), an atlas vertebra fragment (804. 13-1) and a proximal scsamoid (802. 8-1). 15. 2. 3. 28 Gazella sp. (Plates 15. 9-10)
Gazelle bones are common among the Douara Cave animal bones (Table 15. 11). The horncores are mainly broken; of seven, four are male and three female. The only fairly complete male horncore is illustrated in Plate 15. 9. In side view, it is relatively curved; in front view, it shows no torsion. It is moderately compressed: its base is broken, but just above the break it measures ca. 29.5×ca. 20.5 mm; its preserved length is ca. 150 mm along the anterior edge. A broken tip (1005. 13-1) appears less compressed. The only fairly complete female horncore is also illustrated (Plate 15. 9); it is slightly curved backward, measuring (12.5)×(11.3) mm at the base; its preserved length is ca. 65 mm along the anterior edge. Teeth and postcranial bones are fairly small, as the following measurements show:
Harrison (1968) recognizes three living species of gazelle in the Near East: G. gazella, the smaller G. dorcas, and G. subgutturosa. He regards G. marica, described by Thomas from Central Arabia, as a subspecies of G. subgutturosa (Groves and Harrison, 1967). All three species overlap widely in distribution; the features that best distinguish between them-such as pelage, the insertion and shape of the nasal bones, and the spacing between the horncore bases in the male- are not commonly preserved in archeological samples. There are differences in horn shape, but intraspecific variation is considerable. Gasella is abundantly represented in many assemblages from Near Eastern cave sites, but little progress has been made with specific identification. Several extinct species have been described, but apparently with little justification, as intraspecific variation in horn shape has been underestimated. Identification is complicated by the possibility that more than one species may be represented in a sample; and while there are size differences, G, dorcas being smaller than G. gasella, and G. subgutturosa slightly larger and more strongly built, these differences do not appear to be large enough to give any clear distinction (compare measurements in Harrison, 1968; also in Davis, 1974). Identification of the Douara Cave Gazella is thus problematic, especially as more than one species may be represented. The small size of the bones and the slenderness of the one complete metacarpal suggest G. dorcas or G, gazella rather than G. subgutturosa; the curvature of the male horncore and the shortness of the female horncore suggest G.gasella rather than G. dorcas. The female horncores are most unlikely to be G. subgutturosa, as females of that species normally either are hornless or carry fairly large horns (G. s. marica). But closer identification must await a detailed study of osteological distinctions between the species-and especially of postcranial distinctions. 15. 2. 3. 29 Incertae sedis A tooth fragment with rugose enamel (802.6-7) appears to be the posterior part of a moderately hypsodont M3, of about the size of Gazella. Its shape resembles Gazella: Capreolus is more brachydont and has a marked basal swelling, with small basal pillars, both of which are absent in this specimen; but the enamel in Gazella is normally not rugose. 15. 2. 3. 30 Caprini: Ovis and cf. Capra (Plates 15. 11-12)
Caprine bones are even more abundant in the Douara Cave assemblage than are those of Gazella (Table 15.11). Distinction between the different caprine genera is far from simple. Criteria for separating Capra and Ovis are described by Boessneck, Müller and Teichert (1964), but these are based on domestic stock and can only be applied with caution to the different wild species in these genera. Hemitragus occurs today in Oman, and has been identified from the Pleistocene of Europe, and so may occur on Near Eastern sites; Ammotragus is another possibility that should not be excluded: it is widely distributed across North Africa, and has been reported from numerous North African Pleistocene sites. Capra has been found at a number of Upper Pleistocene sites in the Levant. Capra aegagrus is common at Ksar'Akil (Hooijer, 1961), and C. ibex has been identified from a number of sites, generally in associations suggesting drier conditions, as at Zuttiyeh (Bate, 1927b: Fig. 10, called C. primigenia). It has generally been assumed that all caprines from cave sites in the Levant are Capra, but Ovis has recently been found at G VIII, an Epi-Paleolithic site in the northern Negev (I am indebted to Simon Davis for sending me casts of specimens), and is well known from cave sites in Iran and Iraq. Horncore fragments from Douara Cave are small, and they are not very diagnostic. Several have the surface and texture found in Capra; one of these (1003. 8-2) has an anterior edge that is neither sharply keeled nor broadly rounded and is probably best described as bluntly angled. Another small fragment (802. 1-2) comes from near the tip, and is sharply keeled; but is from a surface unit. Only a small number of the teeth and postcranial bones give any useful indication; while there are too few of these to talk about the material falling into two groups, some agree well with modern Ovis, and other agree with Capra. (Ammotragus and Hemitragus tend to resemble Capra rather than Ovis; criteria for distinguishing Capra, Ammotragus, and Hemitragus by the sort of bone fragment that commonly occurs on archeological sites are still essentially unexplored, and postcranial comparative material is scarce even in major collection.) The following specimens are referred to genus-often, as indicated, with some doubt:
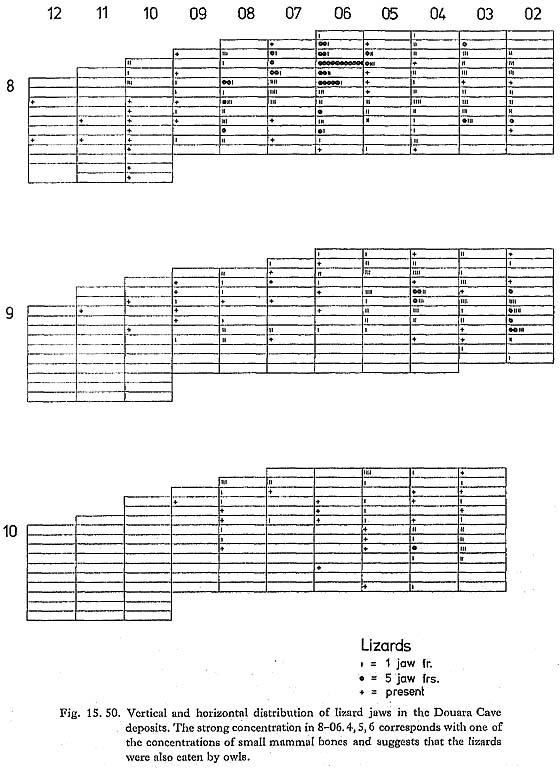
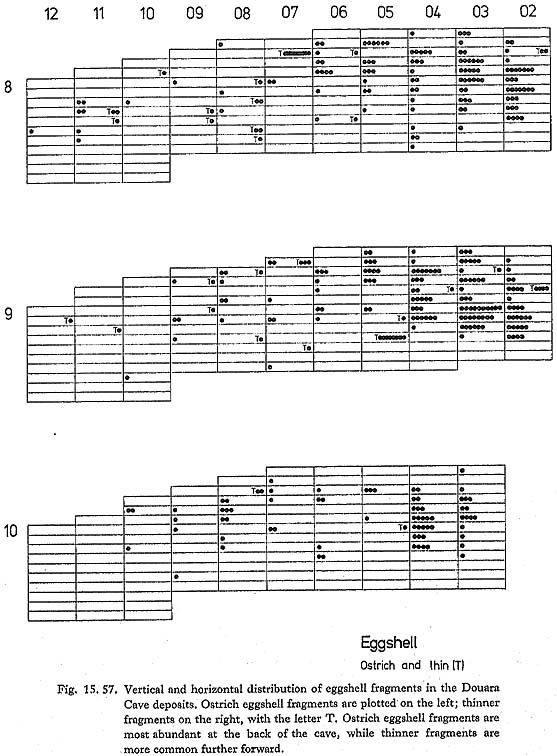
The presence of Ovis at Douara Cave can be taken as reasonably sure, which is not too surprising, especially in view of the recent find from the Negev (see above), the frequency of Ovis (without Capra) at Tell Mureybit, taken to be wild by Ducos (1978), and the recent discovery of wild sheep in Oman (Harrison, 1972). The presence of Capra is more questionable in the absence of reliable criteria separating Capra from Ammotragus and Hemitragus, but the specimens are probably best referred provisionally to Capra. None of the measurements are especially large, and some are fairly small. 15. 2. 4 Other vertebratesA wide range of other vertebrate bones was found at Douara Cave. Bird bones are abundant; most are from fairly small birds, but some large birds are also represented. There are also some ostrich eggshell fragments and some fragments of smaller, thinner eggs. Small lizard bones are also abundant; there are a few snake vertebrae and some tortoise bone fragments. Two fish teeth were found-one is a shark tooth, and probably fossil, the other a sparid or labrid tooth from the front of the cave (10-12. 8). There were also a few fragments of amphibian bones. Some details are given below (Figs. 15. 50, 51 and 57); detailed identification and reports await specialist examination.
|