1.貝類の形態と分類
| ( 1 / 4 ) |
貝類は軟体動物(Mollusca)と呼ばれる動物門に属します。しかし、貝類=軟体動物ではありません。軟体動物でありながら「貝類」ではない分類群も存在するからです。「貝類」とは貝殻(shell)を持つ軟体動物のことを指します。殻の無い分類群と対比させる場合には貝類は有殻軟体動物(shell-bearing molluscs)と呼ばれます。 軟体動物の体制を他の生物と比較してみます。すると軟体動物の体づくりの特徴が明らかになります。軟体動物の基本構造は「多細胞、無脊椎、左右相称、三胚葉性、前口動物、真体腔、無体節性」によって特徴づけられます。 (1)多細胞の体をもつ動物は後生動物(Metazoa)と呼ばれます。単細胞動物または原生動物の対語になります。軟体動物はもちろん多細胞動物です。 (2)軟体動物の体内には脊椎骨がありません。従って、便宜的に「無脊椎動物」に分類されます。 (3)動物の体の相称には放射相称、二放射相称、左右相称などが区別されます。軟体動物は基本的には左右相称(bilateral symmetry)です。腹足類のように左右非相称になる分類群も多数存在しますので全てに当てはまる特徴ではありませんが、基本形は左右対称と考えられます。 (4)発生の過程では軟体動物の体の組織は、内胚葉、中胚葉、外胚葉の3種類の細胞の層(胚葉germ layer)から形成されます。そのような動物群は三胚葉動物(Triploblastica)と呼ばれ、大部分の後生動物は三胚葉動物に含まれます。 (5)軟体動物の成体の口は、卵の表面の細胞層が陥入してできる原口(blastopore)に由来します。原口がそのまま成体の口になる動物は前口動物(Protostomia)と呼ばれます。一方、原口とは独立に口が形成される動物は後口動物(Deuterostomia)と呼ばれます。 (6)体腔(body cavity、coelom)とは体壁と内臓との間の空所のことを指します。体腔は発生的な出来方の違いから原体腔(protocoel)(=無体腔+擬体腔)と真体腔(deuterocoel)が区別されており、中胚葉由来の細胞層に裏打ちされているものが真体腔、されていないものが原体腔です。軟体動物の場合、心臓を取り囲む囲心嚢(pericardium)、排出器官の内腔、生殖器官の内腔が真体腔を構成しています。すなわち、軟体動物の真体腔は尿と生殖物質の形成の機能を担っています。 (7)軟体動物には体節(segment)がありません。内部には一見して体節のように見える繰り返し構造(serial repetition)を持つ分類群もあります。例えば、無板類、多板類、単板類には、神経系、筋肉系、鰓、排出器官、生殖巣、心房の配置に体節状の構造が認められるものがあります。ところが、これらの器官の数と配置は相互に関係がなく、節足動物や環形動物に見られるような規則的な体節とは異なっています。また、外見上でも分節構造を欠くという点で異なります。 以上は軟体動物の基本的な体の構造を説明する概念ですが、これだけでは軟体動物の特徴を表してはいません。以下にそれぞれの器官に注目しながら軟体動物の一般的な特徴を説明したいと思います。「貝殻、外套膜、外套腔、櫛鰓、歯舌」などが重要なキーワードになります。 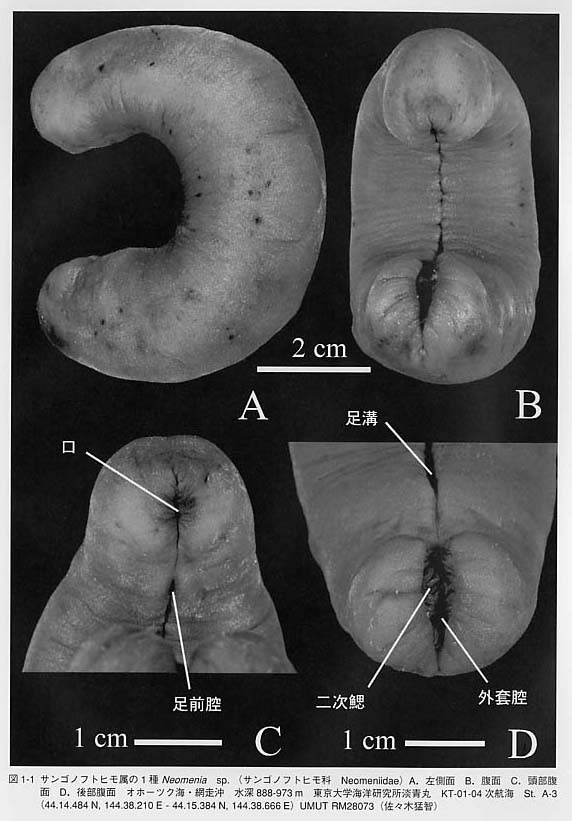 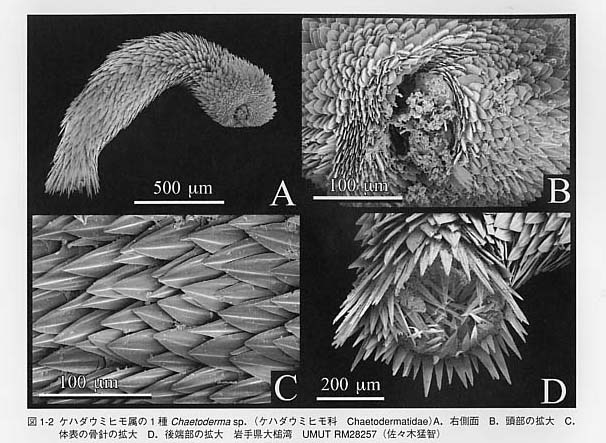 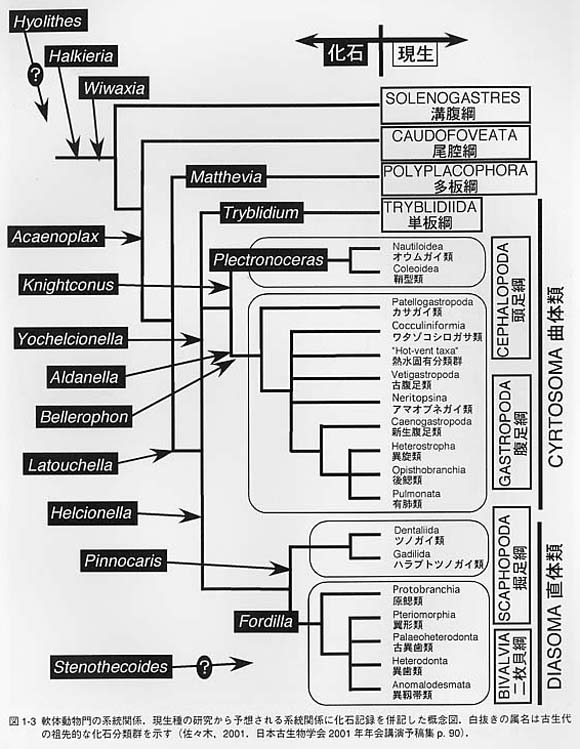 しかし、溝腹類と尾腔類の両者の間には主に5つの大きな違いが認められます。(1)腹溝溝腹類には腹側に腹溝=足溝(ventral groove)がありますが、尾腔類にはありません。(2)体形尾腔類では体の前後にくびれを生じ、頭部が膨らみ尾部が末広がりになる傾向があります。溝腹類は滑らかな紐形の体を持ちます。(3)消化管溝腹類は単純で真っ直ぐな消化管を持ちますが、尾腔類では中腸の腹側に消化腺が分化します。(4)生殖溝腹類は雌雄同体ですが、尾腔類は雌雄異体です。(5)鰓尾腔類には1対の二葉型(bipectinate)の櫛鰓が存在します。溝腹類には鰓を欠きますが、二次的にひだ状あるいは乳頭状の呼吸構造を持つことがあります(図1-1D)。 無板類は全て海産で、生息環境は種によって異なります。溝腹類は群体性の刺胞動物にからみついて棲息しています。日本産ではカセミミズEpimenia babiとサンゴノフトヒモNeomenia yamamotoiが比較的よく知られている種です。尾腔類は大部分が1cm程度あるいはそれ以下の小型種です。外套腔を上にして泥底または砂泥底に埋没し、底質中の有機物を食べています。日本産の種ではアッケシケハダウミヒモChaetoderma akkesienseとヤマトケハダウミヒモChaetoderma japonicumが記載されています。 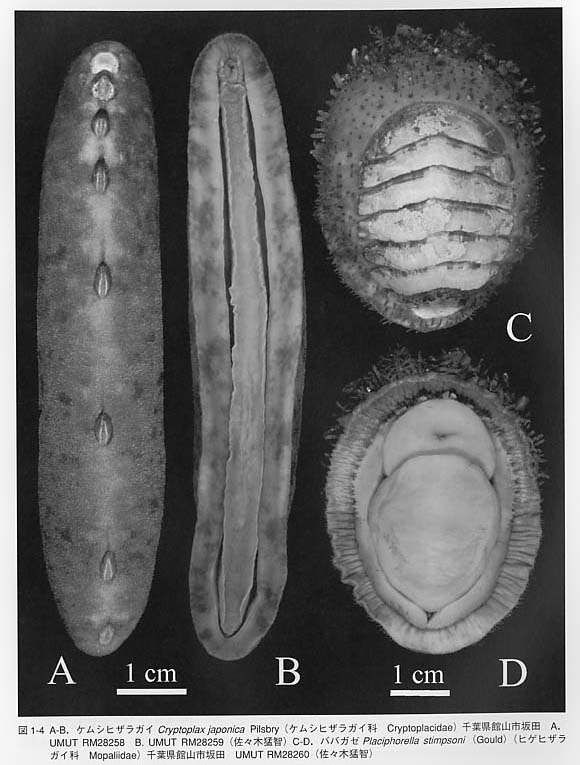
背側には8枚の石灰質の殻板(shell plate、valve)をもちます。しかし、化石の多板類には殻板が7枚しかないものも報告されており、8枚の殻を持つ新多板類NeoIoricataと7枚の殻を持つ古多板類Paleoloricataに区別されています。殻板は体の正中線上に並びます。殻は表層(tegmentum)と連接層(articulamentum)の2層からなります。連接層は前側に突出部を形成し、重なり合います。最も前の殻板は頭板(head valve)、後側の殻は尾板(tail vaIve)、それらの間に位置する殻板は中間板(intermediate valve)と呼ばれます。発生の過程では殻板は最初に7枚が形成され、最後に1枚が追加され8枚になることが知られています。 殻は微細な管状構造によって貫かれています。サイズは一定ではなく、大型のものは大孔(megalopore = macropore)、小型のものは小孔(micropore)と呼ばれます。管状構造の内部には枝状器官(aesthete)があり、大孔・小孔に対応して、大枝状器官(macraesthete)と小枝状器官(micraesthete)と呼ばれます。分類群によっては大枝状器官にレンズを備えた殻眼(shell eye = valve eye)を持つものがあります。 殻の一部または全ては肉帯(girdle)によって覆われています。オオバンヒザラガイCryptochiton stelleri(図1-5)は殻が完全に覆われる例です。肉帯は外套膜が厚く変化したものと考えられています。肉帯の表面には石灰質または角質の棘(spine)、鱗片(scale)、ひげ状突起(bristle)が存在します。 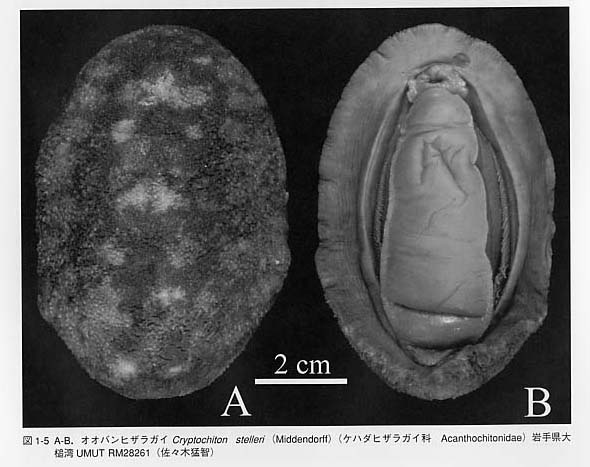
足の周囲には外套溝(pallial groove)が発達します。この溝が他の軟体動物の外套腔に相当します。水を外套溝内に取り入れるときは、肉帯の前の部分を持ち上げます。外套溝には6〜88対の鰓があり、水流は鰓の間を通って後ろへ流れます。鰓の数は体の左右で一致しない種もあります。 歯舌は17列の歯舌歯からなります。そのうちの1対は特に大きく、大側歯と呼ばれます。大側歯の先端は黒く磁鉄鉱(magnetite)で覆われます。この部分は磁性を持っており、磁石に近づければ引き寄せらます。しかし、この磁性のもつ意味は分かっていません。 多板類の消化管の配置は複雑ですが、内部構造は単純です。特に胃の内部には他の軟体動物に見られるような複雑な嚢状の構造が発達しません。腸は長く、腸は前腸(anteriorintestine)と後腸(posterior intestine)が区別され、両者の間には腸弁(intestinalvalve)と呼ばれる弁がある点が特徴的です。 食性は主に藻食ですが、雑食や肉食の種も知られています。肉食性種の代表例はババガゼで、体の前側の肉帯を持ち上げて待機し、小型甲殻類などの餌生物が近づくと肉帯を振り下ろすようにして押さえつけ捕食します。 大部分の多板類は雌雄異体です。しかし、僅かに雌雄同体の種類も知られています。受精は体外受精ですが、外套溝内で保育する種も知られています。 神経系は食道神経環、側神経幹(lateral nerve cord)、足神経幹(pedal nerve cord)およびその横連合からなる原始的なはしご状の構造を持っています。 化石としては多くの種が「単板類」として記載されています。特に、古生代の様々な一枚殻(univalve)型の貝類が単板類と呼ばれています。現生のネオピリナ類を含む狭義の単板類に対しては、混乱を避けるためにMonoplacophoraではなくTryblidiidaという名称が用いられています。この名前は化石属のTryblidiumに由来しています。 殻は単一のカップ状です。殻頂は前側を向き、原殻は左右対称です。殻は外側から、殻皮、稜柱層、真珠層の3層の構造を持ちます。殻頂が前方を向く点では腹足類のカサガイ類Patellogastropodaに類似しますが、単板類が分離した複数の筋肉痕と真珠層を持つのに対して、カサガイ類では単一の馬蹄形の筋肉痕を持ち、真珠層はありません。 単板類の体制上の特徴は、一枚の完全な貝殻が形成されること、多くの器官が複数の対になること、原始的なはしご状の神経系を持つこと、の3点に要約されます。 単板類の鰓、収足筋、排出器官の数は分類群によって異なり3〜8対に変化します。ま た、心房や生殖巣も2対あります。これらのくりかえし構造は体節構造の名残と解釈する見方もありましたが、各器官の位置関係が対応しないことから、体節構造とは見なされていません。 神経系は側神経幹と足神経幹が横連絡によって結ばれるはしご状の構造を持つ点では無板類、多板類などと共通です。そのため、かつてはこれらの三者を双神経類(Amphineura)として分類されていました。しかし、ネオピリナ類は以下の点で、無板類、多板類以外の軟体動物に類似しています。(1)内臓塊を被う単一の大きな殻を持つ、(2)幼生期に特別な殻(原殻)を形成する、(3)外套膜の縁辺部は3葉に分かれる、(4)外套膜に石灰質の骨針を欠く、(5)殻皮腺(periostracal gland)は外套膜縁の腹側に位置する、(6)口の周辺に触手状の構造を持つ、(7)顎板を持つ、(8)胃に晶体(crystalline style)を持つ、(9)側神経幹の後端が無板類・多板類では直腸上横連絡(suprarectal commissure)であるのに対し、ネオピリナ類では直腸下横連合(subrectal commissure)を持つ、(10)平衡胞を持つ。従って、双神経類は神経系以外の形質では支持されないことになり、現在では、単板類・頭足類・腹足類・二枚貝類・掘足類に対して貝殻亜門 (Conchifera)という分類群が用いられています。一方、無板類と多板類に対しては石灰質の骨針を共有するという特徴からAculiferaという名前が与えられています。 |
| 前頁へ | 表紙に戻る | 次頁へ |