





Lespedeza homolobaNakai
被子植物}メ科
徳島県名東郡加茂名村
二階重樓
1912年10月8日採集
選定基準標本(レクトタイプ)
総合研究資料館、植物部門
 | 16-1 ツクシハギ(Lespedeza homoloba Nakai) |
日本産のハギ属の研究は、1867年ミケエル(Miquel)に始まるといってよい。オランダの植物学者ミケエルは、シーボルト(Siebold)やビュエルガー(Buerger)が日本などで採集した標本に基づいて日本に11種のハギ属の種が産することを報告している。この研究で、マルバハギ(L. cyrtobotrya)、キハギ(L. buergeri)などが新種として発表された。現在これらの標本はオランダのライデン植物標本館に所蔵されている(挿図2)。筆者も研究に際してこれらの標本を利用させていただいた。その後の1873年にロシアのアカデミー会員である、マキシモヴィッチ(Maximowicz)によりハギ属の分類大綱の骨子がつくられた。マキシモヴィッチはハギ属を3亜属に分けた。そのうちsubgen. Campylotropisと subgen. Microlespedezaは今日のハナハギ属(Campylotorpis)とヤハズソウ属 (Kummerowia) であり、現在のマメ科植物の研究者はそれらを別属として扱っている。マキシモヴィッチは、今日のハギ属に属する植物をすべてハギ亜属(subgen.Lespedeza)に分類した。ハギ亜属はさらに2つのサブグループ、すなわちヤマハギ節(sect.Macrolespedeza)とメドハギ節 (sect.Lesped�eza)に区分される。
 | 16-2 シーボルト採集のL. buergeri(上)とビュルガー採集のL. cyrtobotrya(下)。オランダのライデン植物標本館所蔵。 |
ある種をその類似種から区別する際、非遺伝的な形質の差異を取り上げるのは適切ではない。多くの形質の変異性を知り、その中から個体群ないしは種として安定的な形質を見いだすことは、あらゆる生物の分類群認識に欠かせない。
ところで、原則として個体ごとに遺伝子は異なるので、個体性の明瞭な高等植物では、遺伝的な差異のまったくない非遺伝的変異のみを調べることは容易ではないと考えられる。しかし、植物は一個体に同じ部分を多数つくるものが多く、非遺伝的変異を調べることはそんなに困難なことではない。
そこで、筆者は、ハギ属ヤマハギ節の種を認識するために形質の変異性を調べた(Akiyama 1988)。方法としては、遺伝的に均一と考えられる一個体に咲く花の形態を調べ、形質の可塑性(phenetic plasticity)を明らかにした。その結果により、一個体内で形質の現れに差異がみられる形質と、個体として安定している形質を探し出した。花の形態は生長する過程で変わる。つまり、蕾から開花時の花に至る過程で、花の各部分の相対生長は異なり、花弁の形態などに変化がみとめられる(挿図3)。そこで、比較のためには、一個体から調べる花は同じ生長段階であるものでなければならない。花としての機能を果している開花直後の花が目的に適していると判断されたので、一個体から得た多数の開花直後の花を双眼実体顕微鏡下で解剖し、描画装置を用いて花の各部の形態を調べた。
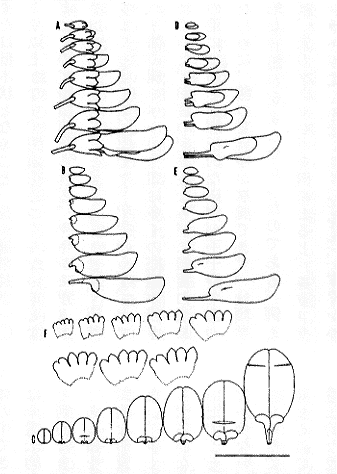 | 16-3 一個体から得た生長段階の異なる花。A:花の全形(横から見た図)、B:旗弁(横から見た図)、C:旗弁(広げた図)、D:翼弁と龍骨弁、E:龍骨弁、F:萼(広げた図)。横線は1cmを示す(以後同様)。 |
可塑性の例として、Lespedeza homoloba の原記載によく合致する個体の花を図示する(挿図4)。長さは11.0〜13.9ミリであり、細部の形にも変化がみられる。爪部の長さの程度には差がみられるが、明らかな爪部をもつ。内側に折りたたまれている耳状突起が腎形でよく発達している。旗弁の先端の形は微凹形である。これらの点は、調べた花の間では変わらず、個体として安定している形質の現れと考えられる。
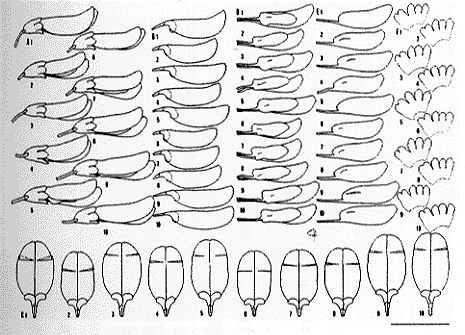 | 16-4 一個体から得た10個の花の変異(記号は挿図3に同じ)。 |
1つの個体群をつくるすべての個体の花を採取し、解剖し、図化し、計測した(挿図5)。個体としては安定していた形質の中には、個体群としても安定しているものと個体群内では変化が大きくなるものとがある。先に記した旗弁の先端の形は、一個体群内では微凹形で微突頭のものがみられ、個体群内では安定していない。明らかな爪部をもつ点と耳状突起が腎形でよく発達する点は個体群内でも安定している。個体群内でも安定している形質は、その形質においてある個体群の集合と他の個体群の集合との間に明瞭な差異が認められれば、それぞれの個体群の集合を種とし、その際の識別に用いられた形質を指標形質とすることができる。
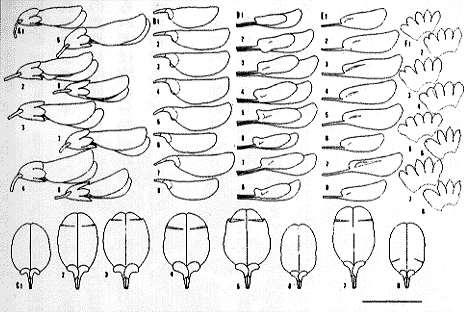 | 16-5 一個体から得た花の変異(記号は挿図3に同じ)。 |
各個体群における形質を調べた結果(挿図6)、それぞれ種として記載された、先に述べたツクシハギ(L. retusa)、ニッコウシラハギ(L. nikkoensis)、ハナキハギ(L. rotundiloba)、センダイハギ(L. sendaica)の各種はツクシハギ(L. homoloba)と同種と判断した。L. homolobaに対して、はじめヤブハギという和名が中井により与えられたが、同じマメ科のヌスビトハギ属のDesmodium podocarpumにもこの和名が用いられる、すなわち、この和名には同名異物のものがあり、これを用いることは情報混乱の原因となるので、現在ツクシハギの和名を用いる理由はここにある。
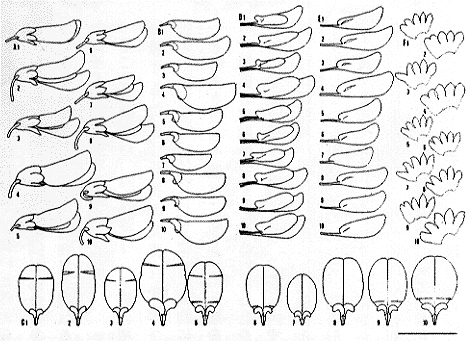 | 16-6 日本各地から得た花の変異(記号は挿図3に同じ)。 |
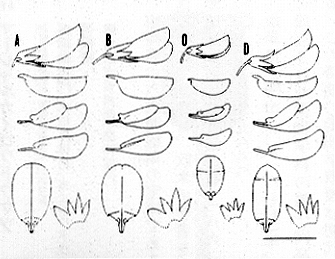
このような解析を行った結果、ヤマハギ節には、ヤマハギ(L.bicolorTurcz.)、クロバナキハギ(L.melananthaNakai)、マルバハギ(L.cyrtobotryaMiq.)、ツクシハギ(L.homolobaNakai)、タイワンハギ(L.formosa(Vogel)Koehne)、ケハギ(L.patensNakai)、オクシモハギ(L.davidiiFranch.)、キハギ(L.buergeriMiq.)、チョウセンキハギ(L.maximowicziiC. K. Schneid.)の九種があることが判明した。各種の相違点は挿図7に示した。
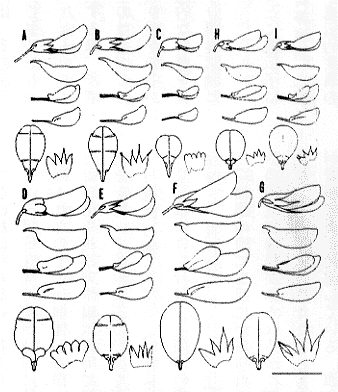 | 16-7 ヤマハギ節各種の花(A: L. bicolor Turcz.、B: L. cyrtobotrya Miq.、C: L. melanantha Nakai、D: L. homoloba Nakai、E: L. formosa(Vogel)Koehnesubsp. velutina(Nakai)S. AkiyamaetH. Ohba、F: L. patens Nakai、G: L. davidii Franch.、H: L. buergeri Miq.、I: L. maximowiczii C. K. Schneid.)。 |
以下にヤマハギ節の種について解説する。
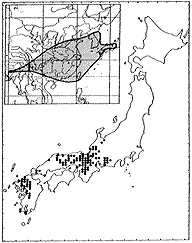
ヤマハギ Lespedeza bicolorTurcz.
異名 L.setilobaNakai, L.spicataNakai et F. Maek.,L.tobaeH.Koidz., L.ionocalyxNakai
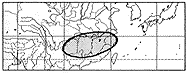
日本全土、朝鮮、中国(東北部)、東シベリアに分布する(挿図8)。日当りのよい場所に生育する。高さ2メートルくらいになり、枝はよく分枝する。花は7〜9月に咲き、明るい紅紫色で、長さ10ミリ前後である。萼は長さ3ミリくらいで、裂片の先端は鈍形または鋭形である。旗弁は翼弁や龍骨弁より長い。旗弁は倒卵形で、基部は次第に細まり、耳状突起は小さい。翼弁は龍骨弁より明らかに短い。果実はほぼ円形または倒卵形で、長さ5〜7ミリである。
日本、朝鮮に分布し、枝の分岐がやや少なく花序が長く、花も大きい個体はヤマハギ(var.japonicaNakai)として、変種として、母変種であるエゾヤマハギ(var.bicolor)から区別されたが、分岐の程度、花序の長さ、花の大きさは連続的に変化し、他に異なる形質がないため、種内分類群として区別することはできない。チャボヤマハギ(var.nanaNakai)は北海道アポイ岳にのみ分布が限られる。高さ30センチ前後の矮性低木であり、ヤマハギの中では例外的に翼弁が龍骨弁より長い。この翼弁が龍骨弁より長いという形質はヤマハギからマルバハギを識別する形質の一つである。分布の限定されたチャボヤマハギと広範囲に分布するヤマハギ(var.bicolor)およびマルバハギとの関係は今後さらに詳細な検討が必要である。
マルバハギ Lespedeza cyrtobotryaMiq.
異名 Campylotropis virgataMiq., L.anthobotryaRicker, L.kawachianaNakai, L.patensNakai var.rotundifolia Honda, L.nakaiiT.Lee
日本では本州(東北地方日本海側を除く)、四国、九州に生育し、朝鮮、中国にも分布する(挿図9)。日当りのよい路傍や沿海地には特に多い。高さ2メートルくらいになり、枝をよく分枝する。花はヤマハギと同様に7〜9月に咲き、明るい紅紫色で、長さ10ミリ前後である。花序は花序軸がほとんど伸長せず、密に花が着き、ふつう葉より短い。萼は長さ4〜6ミリで、裂片の先は鋭尖形である。旗弁は翼弁や龍骨弁より長い。旗弁の形態はヤマハギに似るが、翼弁は龍骨弁より明らかに長く、容易に識別される。茎や枝の毛が開出する個体はカワチハギ(f.kawachiana(Nakai)Hatus.)として区別される。
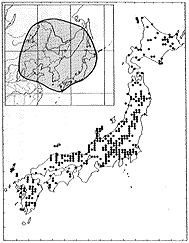 | 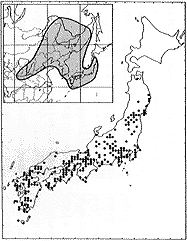 |
| 16-8 L. bicolor Turcz. の分布。 | 16-9 |
クロバナキハギ Lespedeza melananthaNakai
異名 L.homolobaNakai var.higoensisT.Shimizu
分布域がごく限られていて、日本では愛知・熊本および対馬に分布する。朝鮮にも分布するが、やはり分布域は限定されている(挿図10)。ヤマハギに似るが全体に小型である。花はふつう暗赤色であるが、中には花の色が淡く、淡紅紫色のものもある。翼弁は龍骨弁より長いこと、萼裂片は半円形であることからヤマハギと区別される。全体的に繊細な植物であり、花序にはまばらに花が着き、花数も少なく2〜4(まれに6)個である。これらの形質の現れによりマルバハギからも明瞭に識別される。クロバナキハギという和名であるが、キハギに近縁なものではない。
ツクシハギ Lespedeza homolobaNakai
異名 L.nikkoensisNakai, L.retusaNakai,L.rotundilobaNakai, L.sendaicaNakai
前述したように日本固有種であり、東北地方(太平洋側)から九州にかけて低地から低山に分布している(挿図11)。房総、三浦、伊豆半島ではまれで、日本海側の多雪地帯にはほとんど分布しない。キハギに次ぎ木質化がすすみ、またキハギを除いた他の種と比較して耐陰性が強い。葉の上面は中肋を除き無毛で、小葉の先端はふつう鈍形または凹形である。萼裂片は萼筒とほぼ同長またはふつう短く、五裂片がほぼ同形で、先端は鈍形または鋭形であるが、変異が大きい。花はヤマハギより白っぽくみえ淡紅紫色で、長さ10ミリ前後である。旗弁はふつう龍骨弁より短く、耳状突起はよく発達し腎形である。翼弁は龍骨弁より短く、明るい紅紫色である。
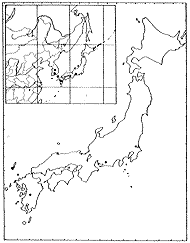 |  |
| 16-10 L. melanantha Nakaiの分布 | 16-11 L. homoloba Nakaiの分布 |
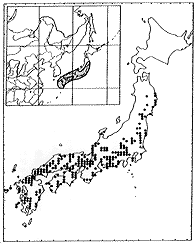
タイワンハギ Lespedeza formosa(Vogel)Koehne
異名 Desmodium formosumVogel, L.viatorum Champ. ex Benth., L.pubescensHayata, L.wilfordiiRicher, L.hayataeHatus., L.ellipticaBenth. ex Maxim., L.bicolorvar. velutinaNakai, L.kiusianaNakai, L.maritimaNakai,L.japonicaL. H. Baileyvar. intermedia Nakai, var. angustifolia(Nakai)Nakai, var.gracilisNakai, var.retusaNakai, var.spicataNakai, L.uekiiNakai, L.tetralobaNakai, L.satsumensisNakai
日本では本州中部以西、朝鮮、台湾、ホンコン、中国、インド(アッサム)に分布する(挿図12)。母亜種(subsp.formosa)はホンコンがタイプロカリテーであり、台湾、中国東部に分布する。中国西南部からアッサムにかけては subsp. elliptica(Benth. ex Maxim.) S. AkiyamaetH.Ohbaが分布し、 これがヤマハギ節の分布の西限である。日本、朝鮮のものはビッチュウヤマハギ(subsp.velutina(Nakai)S. Akiyamaet H.Ohba)である。 明るい場所に生育し、高さ2メートルくらいである。葉の上面はふつう伏した毛が密生する。花は紅紫色で、やや大きく、長さ12ミリ前後である。萼裂片は萼筒と同長か長く、先端は鋭形または鈍形である。旗弁は龍骨弁よりやや長いかまたはほぼ同長で、耳状突起は発達せず狭三日月形〜三日月形である。翼弁は龍骨弁より短い。九州南部薩摩半島と男女列島にはサツマハギ(またはナンゴクチョウセンヤマハギ var.satsumensis(Nakai)S. Akiyamaet H.Ohba)が分布する。サツマハギは、ふつう植物体が矮性であり地際から多数の枝を叢生し枝の毛は開出する。
ケハギ Lespedeza patensNakai
異名 L.bicolorTurcz.f.sericeaMatsum., L.grandifloraH.Koidz., L.formosa(Vogel)Koehnef.sericeaHatus., L.thunbergii(DC.)Nakai f.sericeaHatus.,L.cyrtobotryaMiq. var.macranthaHonda, L.grandisKoidz.
日本固有種であり、本州の日本海側の多雪地に分布する(挿図13)。日当りのよい土手や河原に多くみられる。地上茎および枝は一年生であり、毎年地際の茎から多数の枝を叢生する。枝はよく伸長し、長さ1メートル以上となる。開花時には垂れるものもある。葉の上面は中肋を除き無毛または時に有毛である。開花はふつう晩夏から初秋であるが、初夏から咲き始めるものもある。花はヤマハギ節の中で最大であり、15ミリくらいになる。旗弁は龍骨弁より明らかに短く、爪部はごく短い。翼弁は倒卵形で龍骨弁の半分くらいの長さで、開花時に斜上する。
 | 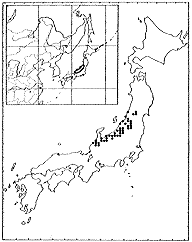 |
| 16-12 L. formosa (Vogel) Koehneの分布 | 16-13 L. patens Nakaiの分布 |
オクシモハギ Lespedeza davidiiFranch.
中国中部に分布する(挿図14)。日本には自生しない。枝には稜があり、小葉は長さ8センチ、幅5センチに達し、ヤマハギ節の中で最大である。葉の上面は伏した毛が密生する。花は紅紫色で、長さ12ミリ前後である。萼裂片は三角形、先端は鋭尖形である。旗弁は龍骨弁とほぼ同長であり、翼弁は最も短い。本種は本学名誉教授故前川文夫により、中国から日本にもたらされ、1966年オクシモハギとの和名がつけられた。資料館には中国で前川が1941年に採集した標本などが収蔵されている。
 | 16-14 L. davidii Franch.の分布 |
キハギ Lespedeza buergeri Miq.
異名 L. oldhamii Miq.
本州(東北地方および中部地方の日本海側を除く)、四国、九州、中国に分布する(挿図15)。耐陰性が強く、林の中にも生育する。また最もよく木質化する。冬芽の鱗片葉および葉は2列互生に位置し、上記の各種からは容易に識別できる。花は初夏から初秋に咲く。花は他の種よりやや小型である。萼裂片の先端は鈍形または鋭形であるが、鋭尖形とはならない。萼の基部の小苞は円形から広卵形である。旗弁は黄色で基部付近に紫斑があり、龍骨弁より短く、さらに翼弁よりやや短いかほぼ同長である。耳状突起はよく発達する。翼弁は紅紫色または紫色である。龍骨弁は淡黄色である。
チョウセンキハギ Lespedeza maximowicziiC. K. Schneid.
異名 L.buergeriMiq.var.praecoxNakai, L.friebeanaSchindl., L.densifloraUyeki, L.tomentellaNakai, L.oldhamiMiq. var.tricolorNakai
対馬と朝鮮南部および中国東部に分布する(挿図16)。キハギ同様によく木質化し、同様の葉序をもつが、花は紅紫色で、萼裂片先端は鋭尖形である。花は早いものでは五月下旬頃に咲き始める。旗弁は紅紫色で、翼弁より長い。龍骨弁は淡紅紫色で旗弁より長い。
 |  |
| 16-15 L. buergeri Miq. の分布 | 16-16 L. maximowiczii C. K. Schneid. の分布 |
前述の九種の中では、キハギは、葉序、花の色などの形質で他の種と著しく異なるため、キハギが一方の親となって生じた雑種は例外的に識別が比較的容易である。
キハギとマルバハギが混生する集団では、雑種がしばしば見出される(Akiyama&Ohba 1982)。雑種個体は、オクタマハギ(L.×cyrto-buergeriS. Akiyama et H. Ohba)と名づけたが、花の形が中間的になるだけでなく、葉形、葉序などの栄養器官の形質も中間的な現れとなっている。花粉の染色率はキハギとマルバハギでは94.4〜100パーセントであるのに対して、オクタマハギでは染色率は10.4〜75.5パーセントで、かつ変形してつぶれている花粉が多く観察された。花粉の稔性は、両親種よりも低下していると考えられる。オクタマハギは、両種が同所的に分布する各地で個体数は少ないもののしばしば見出されている。ハギ類は総じて日当りのよい場所に生育する。崖崩れ、伐採等により日当りのよい場所ができると、マルバハギやキハギなどが生える。キハギの開花時期は初夏から初秋にかけてであり、一度に開花するというよりもむしろ少しずつ咲き続ける。マルバハギは初秋に一度に開花する。この時期に両種の雑種が形成されると考えられる。その後、他の樹木が生長するにつれて、日当りが悪くなってくる。するとまずマルバハギの生育が難しくなる。マルバハギの花つきは少なくなり、もはや雑種は生じなくなる。一方キハギは比較的耐陰性があるために、以前と同様に生育を続ける。両種の雑種個体もしばらくは生育を続ける。その後やがてマルバハギは生育できなくなり、キハギと雑種個体のみになり、さらにはキハギのみとなる。したがって、マルバハギとキハギの分布域の重なる所では、しばしば雑種個体が見いだされるものの、その個体数はごく限られており、稔性が低下しているために、その子孫はほとんど生じないと推定される。やがては、雑種個体も消滅する。
ツクシハギとビッチュウヤマハギの間に生じる雑種は、稔性がほとんど低下せずに、次世代を生じ、個体群の中に存続していくと考えられる。両種は、西日本各地でしばしば同所的に生育しているが、花の形態、葉の上面の毛によってふつうは明瞭に識別される。しかし、岡山県で見出された個体群では、様々な形態をした両種の中間形が存在した(Akiyaya&Ohba1983)。花粉染色率は93.0〜98.2パーセントと高く、ほぼ正常の稔性を有すると推定された。花粉の稔性が低下していないことから、これらの個体は、両親種との戻し交雑および雑種個体間での交雑を行っていると推定される。このような雑種個体群は両親種間の識別をも困難にしているほどである。前述のオクタマハギの場合とは異なり、両親種の形質を様々な程度に組み合わせてもつ個体がみられるので、種小名の命名は行わなかった。しかし、雑種個体群が存在するにもかかわらず、雑種個体(群)は全体からみるとごくわずかである。この点から、通常は何らかの生殖的な隔離が存在していると判断される。しかし、完全ではない生殖的隔離がどのように存在し、それぞれの種が独立性を保っているか、非常に興味深い問題である。
ミヤギノハギ(L.thunbergii(DC.)Nakai、挿図17)は、野生種であるケハギによく似ているが、枝はよく伸長して下垂し、葉は深緑色、小葉の先は鋭形、花色は濃紅紫色である。ケハギの形態の変異の幅が広いのに対して、ミヤギノハギは形態の変異の幅が極端に狭く、ごく限られた個体から株分けなどの栄養繁殖の手段により増殖されたと考えられる。ケハギの中の一型から作り出された園芸品であると推定されるが、現在栽培されているものは稔性の低下したものが多くみられることから、ケハギと同種とはせず、園芸種として独立させるのが適切である(Akiyama 1988)。栽培は江戸時代に始まったと推定される。1784年にツュンベルク(Thunberg)が著わした日本の最初の植物誌であるフロラ・ヤポニカ(Flora Japonica)では、本種がリネウスの発表したHedysarum heterocarponの学名で載っている。このH. heterocarponとして載っている植物は、後にまったく別の植物であることが判明した。これを最初に明らかにしたのは、ド・カンドル(de Candolle)であり、彼はこの植物(すなわちミヤギノハギ)をヌスビトハギ属の種として、ツュンベルクに献名しDesmodium thunbergiiと命名した。その後、1927年に中井教授によりハギ属に組替えられた。挿図18は中井教授による、ツュンベルクの採集したミヤギノハギの標本の模写である。ツュンベルクの後に来日したシーボルトも日本のハギに興味をもっていたと思われる。シーボルト自身が日本で採集し、その後その標本に基づいてミケエルによってLespedeza sieboldii と命名された植物は、ミヤギノハギである。また、彼が所蔵していた日本植物の図説が出版された(Kimura&Grubov 1993)が、その中にもミヤギノハギの図があり、その図はライデンの植物標本館に収蔵されるL.sieboldiiの標本とよく一致する。
 | 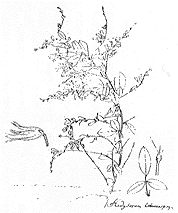 |
| 16-17 A: L. japonicaL. H. Baileycv. NipponicaNakai、B: L. japonicaL. H. Baileycv. japonica、C: L. japonicaL. H. Baileycv. HiratsukaeNakai、D: L. thunbergii(DC.)Nakai | 16-18 中井教授による、ツュンベルクの採集したL. thunbergiiの標本の模写。 |
ニシキハギ(L.japonicaL .H. Baileycv.Nipponica、挿図17)もミヤギノハギと同様 によく栽培されるが、ミヤギノハギとは異なり変異の幅が広い。ニシキハギはビッチュウヤマハギと同種とされることもあるが、その変異は完全には重ならない。前述したようにハギの類では雑種形成がしばしば生じることから、ニシキハギは野生種であるビッチュウヤマハギが栽培されている間に他の種と交雑をして生じたいろいろな型の雑種の集合と考えられる。したがって、これも園芸種として独立させるのが適切であると判断した。
(秋山 忍)
 berLespedeza Michx. und ihre nachsten Verwandten. Engler, Bot. Jahrb.49: pp.570-658.
berLespedeza Michx. und ihre nachsten Verwandten. Engler, Bot. Jahrb.49: pp.570-658.