植物の矮小化 |
|
秋山 忍 国立科学博物館植物研究部 |
|
高山帯に生活の本拠をもつ植物を高山植物というが、一般に高山植物には、比較的小形の多年生草本や小低木が多い。とくに草本植物では背丈のわりに花が大きいものが多いことが知られている。 植物の矮小化は、高山帯に限らず、極地域、乾燥地などでもみられる現象である。矮小化をもたらす環境的要因として、生育可能期間が短いことなどがあげられている。しかし、植物の矮小化には明確な定義はなされておらず、また体系的な認識もされていない。ここでは、これまで東京大学総合研究博物館が中心となって行ってきた調査・研究成果を中心に記す。 ヒマラヤなどの熱帯や亜熱帯にある、低中緯度の高山帯では植物の生長可能な期間が短いだけでなく、風、日射、紫外線が強く、温度や気圧は低く、日変化が大きい。日射が強く、日変化が大きい点などは、夏は白夜となり日較差の小さい極地域とは大きく異なる。高山帯では、しばしば生育期間中も夜間は厳しい放射冷却にさらされ、降霜ときには降雪にさえみまわれることもある。また、気圧が低い点は、高山帯に限られる。 ヒマラヤ高山帯における植物の生長可能な期間は、海抜三九〇〇−四二〇〇メートルでは十五週、五〇〇〇メートルでは十週であると推定されている[Mani 1978]。植物は、この短い期間に、芽を出し、葉を展開し、開花・結実させなければならない。植物体自体が小形化することによって、つまり、生長開始後の比較的早い時期に開花・結実することにより、生長そのものを短い期間で成し遂げることができる。これは、短い生長可能期間しかない環境下における生活に有効であると考えられている。 矮小化には、(一)近縁種と比較してある種が小形な場合と、(二)同種内で小形化する場合がある。高山植物は、ふつう(一)の場合にあたり、近縁種に比較して植物体が小形になっているが、分類群の違いに拘らずに、クッション形やロゼット形などの類似の形態を有する植物が多くみられる。(二)の場合には、一時的な小形化と遺伝的に安定した小形化がみられる。
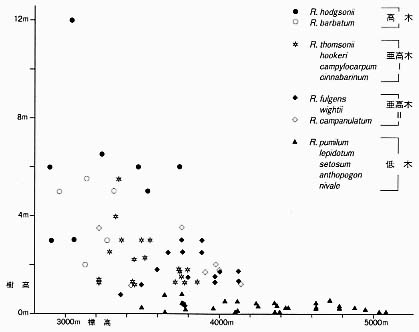
i 木本植物山に登り、標高が高くなるにつれて、ふつうは、高木がなくなり、低木がみられ、やがては、ごく矮小な低木しかみられなくなる。 シャクナゲ属(ツツジ科)は、ヒマラヤで多様化が著しい。日本では、高木になるシャクナゲ属植物はみられないが、ヒマラヤでは、ネパールの国花であるRhododendron arboreum Sm. のように樹高二〇メートルになるものもある。これに対して、高山帯では、小形で、高さわずか一〇センチほどに低木化したR. nivale Hook. f. が生育している。一九八三年以来行われている東京大学を中心としたヒマラヤ地域の植物調査のひとつとして、ネパール東部のバルン谷沿いでのシャクナゲ属植物の樹高と生育標高が調べられた[Noshiro and Suzuki 1989、鈴木一九九二、Noshiro, Suzuki and Ohba 1995]。亜高山帯林内の海抜三〇〇〇メートルくらいでは、高木であるR. barbatum Wall. ex G. DonやR. hodgsonii Hook. f. が生育し、樹高は一〇メートルを超える個体もみられる。標高が高くなるにつれて高山帯潅木帯の亜高木であるR. thomsonii Hook. f.、R. campylocarpum Hook. f.、R. fulgens Hook. f.、R. campanulatum D. Donなどが生育し、海抜四一〇〇メートル付近では樹高は二メートル以下である。さらに標高が高い所には、これらの亜高木性の種は生育していない。そこには、低木であるR. anthopogon D. DonやR. lepidotum Wall. ex G. Donなどが生育し、海抜五〇八〇メートルにまで達している。グラフ[表1]に示されたように、標高とともに、樹高が低くなり、小形化しているのがよくわかる。このように標高が上がるとともに樹高が低くなるシャクナゲ属植物の木材構造が調べられている[Suzuki and Ohba 1988; Noshiro and Ohba 1993; Noshiro, Suzuki and Ohba 1995]。木材構造のうちでも、導管に大きな違いがみられる。導管の太さは、高木では直径が一〇〇ミクロンを超えるのに対して、高さ一メートル以下の低木では直径三〇ミクロン以下である。また、同じ種で、高木から低木へと樹高が低くなるにともなって、導管の直径も順次小さくなっている。この導管の直径の違いが、実際の樹木の生活にどう働いているかはいまだ解明されていないが、導管が細くなると通導効率は下がるが、気泡による導管閉塞による乾燥化にともなう枯死の危険性が減少して凍結や乾燥に強くなることが予想されている。また、道管要素の長さは、生育場所の高度が高くなるに従って短くなる。どのようにしてこのような現象が生じているかについての詳しい解析は、今後の研究がまたれる。
ii 草本植物(一年生草本)同じ属の他の種と較べて著しく小形な種が高山帯に生育するものとして、ツリフネソウ属(ツリフネソウ科)があげられる。ツリフネソウ属は、多年生草本または一年生草本であり、ヒマラヤでは森林帯に多数の種が生育している。高さ三〇−一〇〇センチくらいになる種が多いが、高山帯に生育する一種、Impatiens occultans Hook. f. は、高さ二−一〇センチくらいで花を咲かせ、種子をつくる[挿図1]。小さいものでは、二枚の子葉が展開した後、わずか二、三枚の本葉を展開しただけで、花を咲かせる個体もある。次節で詳しく述べるRhodiola bupleuroides (Wall. ex Hook. f. et Thomson) S. H. Fu の場合と同様に、茎の伸長がわずかであり、幼形成熟を伴う一種の性成熟過程の縮小であるといえる。高山植物は、ふつう草丈に比して花が大きいことが知られているが、このI. occultans Hook. f. では、花も他のツリフネソウ属植物と比較して小さく、長さ〇・五−一センチくらいである[Akiyama, Ohba and Wakabayashi 1991]。
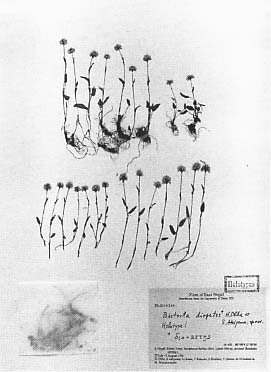
ヒマラヤに分布する他の小形の一年生草本に、地中果をもつLignariella hobsonii (Pearson) Baehni(アブラナ科)[Arai and Ohba, in press]や、繊細な茎をもつKoenigia delicatula (Meisn.) H. Hara(タデ科)などがある。特にチシマミチヤナギ属Koenigia では、K. delicatula やK. nepalensis D. Don などヒマラヤ地域に固有な(ただしK. nepalensis は台湾にも分布するという)数種がある。一方、周極地域からヒマラヤに広く分布する小形のK. islandica L. やヒマラヤに固有であるが多年生草本でマット状に生育するK. nummularifolia (Meisn.) Măsíček & Soják などがあり、ヒマラヤでの矮小化の研究上興味深い。 iii 草本植物(多年生草本)多年生草本にも、一年生草本で記したように、草丈が低く、茎の伸長がわずかであり、幼形成熟を伴う一種の性成熟過程の縮小がみられる。むしろ、高山植物の多くの種は多年生草本であり、一年生の種は少ない。 東京大学のヒマラヤ地域植物調査隊によって、イブキトラノオ属(タデ科)では、高山帯で矮小化したBistorta diopetes H. Ohba & S. AkiyamaやB. sherei H. Ohba & S. Akiyamaが見いだされた[Ohba and Akiyama 1992]。これらの種は、草丈がわずかに五センチくらいで、開花・結実する。 植物体の花茎が伸長せず、地際に花をつける植物があり、ロゼット形といわれる。オオキセワタ属Phlomis(シソ科)[挿図2]、トウヒレン属Saussurea(キク科)など、多くの科にこのように特殊化した種がある。
植物体が小形化し、密集してコケ植物のような生育形態をとった種が、ナデシコ科、ユキノシタ科、サクラソウ科など、直接の系統的な関連のないいくつかの科にみられる。これらをクッション形植物という。 ロゼット形やクッション形は、地表に接していることや密集することで地熱や地中の水分を逃すことなく生長に有効に作用しているといわれている[Ohba 1988]。Androsace tapete Maxim(サクラソウ科)[挿図3]はヒマラヤの高山帯を代表するクッション形植物だが、トチナイソウ属(Androsace)には、このようなクッション形となる種が多い。ナデシコ科ノミノツヅリ属(Arenaria)にもA. densissima Wall. ex Edgew. & Hook. f.やA. glanduligera Edgew. ex Edgew. & Hook. f. などのクッション形植物がある。ユキノシタ科ユキノシタ属(Saxifraga)にもS. andersonii Engl. などが分類されるKabschia節や、S. saginoides Hook. f. & Thomsonなどにクッション形植物がみられる。系統的に関連のない種であるが、植物体のみかけは大変よく似ている[挿図4]。
このようなロゼット形やクッション形のように地表に葉を広げている植物(地表植物または半地下植物とよばれる)は、ヒマラヤに限らず、世界の高山帯と極地域に見られる植物である。挿図5は、アフリカの高山帯でみられる生活形の主なものを示している。ここでも、ロゼット形とクッション形がみられる。また、南アメリカの高山帯でもロゼット形とクッション形がみられる。 実際に高山帯において、ロゼット形およびクッション形の植物の温度と周囲の温度が測定された例は少ない。その一例として、ベネズエラの標高三六〇〇メートルの地点での三日間の測定によれば、Hypochaeris sessiliflora Humb. & al(キク科)という小さなロゼット形植物では、その中心部の最低温度は、周辺の裸地より六度も高い。また、十一回の測定値の平均では、ロゼット中心部の温度は、地表より五〇センチの空気中の温度より、約六度高かった。同じくベネズエラの標高三六〇〇メートルに生育していたクッション形植物であるPlantago rigida(オオバコ科)では、クッションの表面下〇・五センチの所の温度は、周辺の空気中温度より平均で約三度高い[Hedberg and Hedberg 1979]。 このように、実際に、クッション形植物とロゼット形植物では、空気中の温度より高くなっていて、高山における生育期間中の低温に対する適応の一形と考えられている。
i 一時的な矮小化日本の富士山において、匍匐しているカラマツ(マツ科)があることはよく知られている。富士山のお中道沿いには、ハイマツ状となったカラマツが生育し、また、コメツガにも同様な形が知られている[大場一九八五]。このカラマツやコメツガの矮小化は冬の風や雪圧などの環境による一時的な矮小化と考えられる。 ネパール・ヒマラヤでは、ネズミサシ属の一種であるJuniperus recurva Buch.-Ham. ex D. Don(ビャクシン科)は、ネパールでは海抜三三〇〇から四六〇〇メートルの高度に生育する。標高の比較的低い所では、高さ約二〇メートルに達する高木となる。しかし、標高が高くなるにつれ、樹高は低くなり、幹が明らかでなくマット状に広がり、中部ネパールでは海抜四四〇〇メートルにまで生育する。木本植物の小形化では、幹がはっきりしなくなり、茎や枝が地面を這うように広がり、樹形が変わる。これは、温度が低いことだけでなく、風や雪の影響をも強く受けている場合がある。このような樹形が、遺伝的に安定しているか否か、詳細については不明である。矮小化の安定性に関しては、種による違いも考えられる。今後詳しい調査・研究が必要である。 一時的な矮小化が明らかな例として、動物の影響による矮小化がある。ヒマラヤの高山帯では夏に、家畜の放牧が行われている。草食獣の採食が行われている短草型群落では、Bistorta macrophylla (D. Don) Soják(タデ科)やPotentilla griffithii Hook. f(バラ科)などの植物は、草丈が短く矮小である。しかし、その群落に生育するトゲ植物などの低木や多年生草本の防衛植物(トゲがあったり、有毒成分を含んでいて、動物が採食しない)の陰に隠れるようにして生育している個体では、草丈が高く、葉も大きい[高槻一九九三]。これらの種では、矮小化は可逆的な一時的なものである。 ii 安定した矮小化草本植物での小形化は、ふつう茎の伸長が少なく、早くに花をつける。これは、幼形成熟を伴う一種の性成熟過程の縮小で、生長期間が短いことによって引き起こされたと考えられるが、それだけでなく、土壌条件や風などの影響による場合もあると考えられる[大場一九九二]。 高山帯では、植物体の茎がわずかしか伸長せず、花を咲かせる植物がみられる。茎丈は低いが、花の大きさは茎丈が伸長するものとふつうほとんど同じである。生育地の高度の違いに応じて矮小化している植物の例として、Rhodiola bupleuroides (Wall. ex Hook. f. et Thomson. S. H. Fu(ベンケイソウ科)がある。この種はイワベンケイ属の一種であり、ネパール・ヒマラヤからシッキム、ブータン、チベットを経て雲南・四川にまで分布を広げている。ネパールでは海抜二〇〇〇から五六〇〇メートルにかけて割合広く分布しているが、まれには二〇〇〇メートル以下の場所でも見いだされている。この種は、植物体の大きさ、葉の形と大きさの変異が大きい。これまでは、十分な標本資料が蓄積されておらず、これらは一種とされているが、現在、大場・若林による研究が進められている[大場一九八六、および後出「種分化と染色体」の項参照]。海抜二〇〇〇−三六〇〇メートルの林の中に生育する個体は、植物体の高さが二五センチ以上で、葉の長さは三センチ以上である[挿図6a、b]。高山帯下部である海抜三五〇〇・四四〇〇メートルに生育する個体は、植物体の高さがおよそ三〇センチで、葉の長さはおよそ二センチである[挿図6g]。高山帯上部である海抜四六〇〇メートル、四〇〇〇メートル、三六〇〇メートルの地点に生育している個体は最も小形であり、植物体の高さが五センチ以下で、葉の長さは五ミリ前後である[挿図6d-f]。この植物では、矮小化と染色体数の倍数性の間に関連があるので、矮小化は一時的なものではなく、遺伝的に安定したものであると推定される。
Saxifraga pallida Wall. ex Ser.群(ユキノシタ科)は、ヒマラヤから中国にかけて海抜約三〇〇〇メートルより上の高山帯に生育する。海抜が比較的低い所では、枝分かれした花序に花をつける。高い所では、ロゼット上に葉を広げ、その中心部に低い花茎をだし、少数の花をつける。高度の最も低い所の個体は二倍体であるが、高度の高い所の個体は高次倍数体があることが調べられつつある[大場一九八六、若林一九九二]。 以上のように、植物の矮小化には、いろいろな場合がある。また、矮小化は高山という環境への適応の一形であると考えられているが、すべての植物が矮小化するわけではない。矮小化した植物と同じ環境に、対照的な巨大化した植物や特殊な形態をしていない植物も生育するヒマラヤ高山帯は矮小化の研究に最適の研究環境にあるといってもよく、今後の研究成果がまたれる。 |




【参考文献】Akiyama, S., Ohba, H., and Wakabayashi, M. 1991. Taxonomic notes of the East Himalayan species of Impatiens. Studies of Himalayan Impatiens (Balsaminaceae)(1). In: H. Ohba and S.B. Malla Eds., The Himalayan Plants, vol2, pp. 67-89. Tokyo: The Unviersity of Tokyo Press.Arai, K. and H.Ohba, in press. Lignariella hobsonii. Hedberg, I. and Hedberg, O. 1979. Tropical-alpine life-forms of vascular plants. Oikos 33, pp. 297-307. Mani, M. S. 1978, Ecology and phytogeography of high altitude plants of the Northwest Himalaya. London: Chapman and Hall Ltd. Noshiro, S. and Ohba, H. 1993. Altitudal distribution and tree form of Rhododendron in the Jaljale Himal, East Nepal. Journal of Japanese Botany 68, pp. 193-198. Noshiro, S. and Suzuki, M. 7989. Altitudal distribution and tree form of Rhododendron in the Barun Valley, East Nepal, Journal of Phytogeography and Taxonomy 37, pp. 121-127. Noshiro, S., Suzuki, M. and Ohba, H. 1995. Ecological wood anatomy of Nepalese Rhododendron (Ericaceae). 1. Interspecific variation. Journal of Plant Research 198, pp. 1-9. 大場秀章、一九八五年、「三、四 新しい山富士」、山堀越増興・青木淳一編『日本の自然六 日本の生物』、五二−七〇頁、岩波書店 大場秀章、一九八六年、「ヒマラヤ高山帯の植物」『科学』第五六巻三号、一四六−一五二頁 Ohba, H. 1988. The alpine flora of the Nepal Himalayas: an introductory note. In: S. B. Malla and H. Ohba eds., The Himalayan Plants, vol. 1, pp. 19-29. Tokyo: The Unviersity of Tokyo Press. 大場秀章、一九九二年、「七 極限環境での植物の適応」、柴谷篤弘・長野敬・養老孟司編『講座進化七』、二二七−二四五頁、東京大学出版会 Ohba, H. and Akiyama, S. 1992. The alpine flora of the Jaljale Himal, East Nepal, The University Museum, the University of Tokyo. Nature and Culture, No. 4. p. 83 鈴木三男、一九九二年、「ヒマラヤの森林限界・樹木限界」『遺伝』第四六巻九号、一二−一六頁 Suzuki, M. and Ohba, H. 1988. Wood structural diversity among Himalayan Rhododendron. IAWA Bulletin n. s. 9, pp. 317-326. 高槻成紀、一九九三年、「ネパール中部、ランタン谷の草地群落と家畜の影響」『プランタ』第二六号、一〇−一九頁 若林三千男、一九九二年、「高山植物の染色体の特性」『遺伝』第四六巻・九号、三六−四二頁 |
|



| 前頁へ | 表紙に戻る | 次頁へ |