Hiroko Koike
Introduction
There are over 600 known shellmounds of the Jomon period in the area surrounding Tokyo Bay. These shellmounds, or middens, contain many types of food remains that provide useful information on diet and exploitative activities. Jomon culture appears to have been based primarily on hunting and gathering; consequently, environmental factors, especially changes in fauna and flora as food resources, would have had a significant effect on exploitative activities, settlement patterns, and population size.
Seasonality of hunting and gathering activities has been a central subject in the study of exploitative strategy. Since annual layers in the cement of teeth found in excavated mate-rials was first studied by Saxon and Higham (1968), the season during which game mam-mals were hunted has been. estimated by the annulation method (Bourque, 1975; Spiess, 1976; Bourque et al., 1978; Ohyi et al., 1980, 1983).
In Japan, a seasonal-dating project to reconstruct prehistoric exploitation patterns has been working intensively to establish standards of age determination for the sika deer (Ohtaishi, 1978), the wild boar (Hayashi et al., 1977; Nishimura, 1982), two kinds of harbor seal (Naito and Nishiwaki, 1972; Naito and Kano, 1979), the sea urchin (Tald, 1972), and molluscs (Koike, 1980). The project has also discussed the application of the age com-position of excavated game animal populations to the problem of estimating prehistoric hunting pressure.
According to data on living sika deer populations (Ohtaishi, 1978), the age composi-tion of wild deer populations under natural conditions is younger than that of deer populations under protection, the longevity of hunted wild deer being only half the physiological longevity. The age composition of hunted deer populations shows that the per-centage of deer older than five years declines rapidly, and that deer older than eight years are usually rare. These differences in age composition indicate the influence of hunting pressure on wild deer without protection.
Fishery reports of shellfish resources in the tidal flats of the Ariake Sea (Fishery Experi-mental Station of Kumamoto Prefecture, 1951) show that clams from intensively collected areas are smaller than those from protected areas, and that shell height distribution is a useful indicator in the management of shellfish resources.
Hunting pressure estimated from the age composition of an excavated faunal collection can be used as an indicator to infer the intensity of human activity with relation to the animal, or as an indicator of the balance between human consumption and biomass.
Paleobiomass analysis is composed of three steps (Koike, 1983a and see also Fig. 1), The first step is to establish the chronology and distribution of human settlements, thus clarifying the archaeological history of the subject area. The human population, may be estimated from archaeological data on the number and size of settlements. The second step is to reconstruct the prehistoric environment for each archaeological stage in the area. The marine environment, that is, shoreline and tidal flat conditions, is reconstructed by means of paleotemperature, molluscan fossil and micro-fossil analysis. The terrestrial environment is reconstructed by analysis of pollen and plant macrofossils. Paleobiomass can then be estimated from the paleoenvironmental data and modern wildlife management data on living animals.
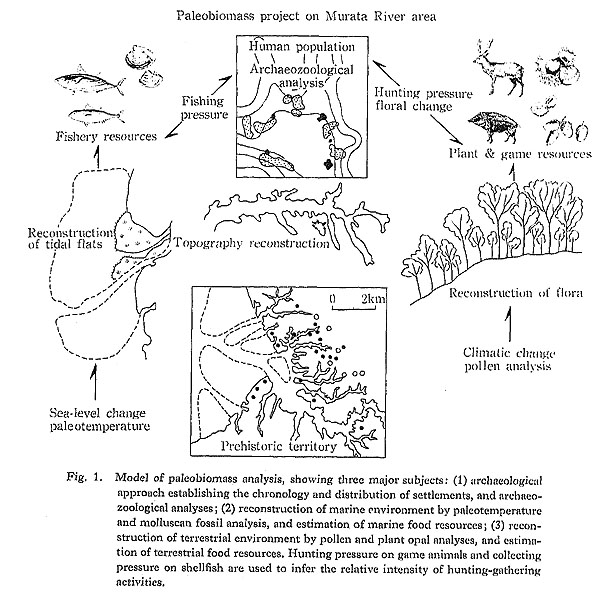
With these data in hand, the third step is to estimate the hunting pressure on game animals and collecting pressure on shellfish. This is done by determining the age composition of archaeologically recovered collections. This paper provides an example of paleobiomass analysis for the Murata River area, where intensive geological and archaeological survey has been prompted by the Chiba Newtown Project.
Archaeological Analysis
Mapping Survey of the Murata River Area
Regions subjected to environmental analysis should include the whole area exploited by the dwellers of a settlement. Hunting of terrestrial game animals and gathering of plant foods in the hilly forest was no doubt usually done over an area of approximately 15-km diameter within which people could forage on foot and still return home the same day. Shells were collected mainly in the tidal flats at river mouths, and fishing was done in shallow water zones along the coast. The Murata River area was selected to serve as a model of paleobiomass analysis because the river system is rather short (spanning 10 km from the head to the recent mouth) and because it flows into Tokyo Bay through large tidal flats at the mouth.
The Murata River is located on the eastern coast of Tokyo Bay, about 8 km south of Chiba City (Fig. 2A). The Murata River system is composed of several branch valleys which separate peninsula-like terraces that rise 20 to 40 m above sea level. A mapping survey of the Murata River area (Education Committee of Chiba City, 1983) showed that large shellmounds are densely distributed on the terraces.
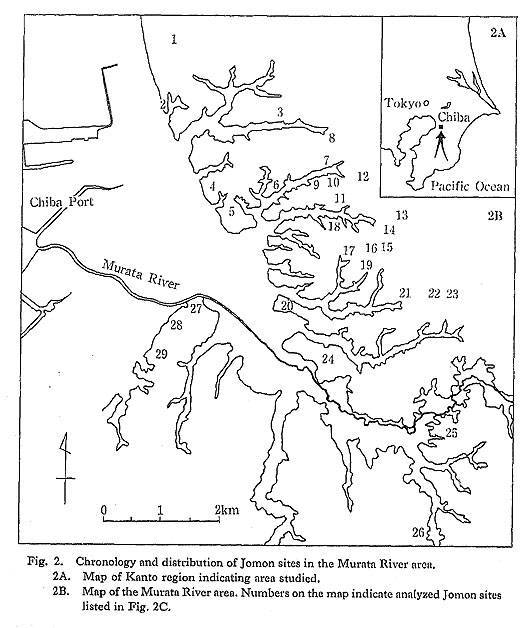
Twenty-nine Jomon sites have been reported so far in the Murata River area (Fig. 2B). Among them, 18 sites are accompanied by sufficient molluscan shell accumulations to be categorized as shcllmound sites. Horseshoe-shaped shell accumulations were recognized at seven sites, while the remaining 11 have relatively small, scattered shell deposits.
The chronology for the Jomon sites of the Murata River area (Fig, 2C) shows that the first Jomon occupation of this area occurred ca. 6000 B.P. during the later stage of the Initial Jomon period (Kayama pottery stage). There were few occupation sites of the Initial and Early Jomon periods, and they contained few artifacts. The number of sites increased during the later stage of the Middle Jomon period (Kasori E pottery stage), and the formation of shell accumulations seems to have been most rapid during the early stage of the Late Jomon period (Horinouchi pottery stage), Some of the shellmound sites continued until the later stage of the Late Jomon period (Angyo I and II pottery stages), until all eventually disappeared in the Final Jomon period (Angyo IIIa etc, pottery stages). The most continuous occupation was at the Rokutsu shellmound site; the largest shell-mound in this area, its occupation continued from Horinouchi to Angyo IIIa times.

Kidosaku Shellmound Site
The Kidosaku shellmound site provides one of the best available samples for paleobiomass analysis, because the entire settlement has been unearthed by the Cultural Property Center of Chiba Prefecture (1979). A total sample of dwelling pits from the terrace, and food remains from the shell layers, offer useful information on population size and data for archaeozoological analysis. Another analytical advantage is that all the potsherds from the site belong to the Horinouchi type, suggesting that the period of occupation there was relatively short.
Population Sise
The entire site, when excavated, revealed 10 dwelling pits, distributed surrounding a central plaza on the very edge of the terrace (Fig. 3A). Each dwelling pit was circular in plan, 4 to 6 m in diameter, with a firepit in the center and postholes along the walls. Three dwelling pits found on the southern fringe of the site (Fig. 3C) had small rounded exten-sions, 2 m wide and 1 m long. Although it is still unclear whether these dwellings with this special feature had a special function (e.g. as ceremonial houses), it is assumed that they were, at the least, important dwelling houses of the settlement.
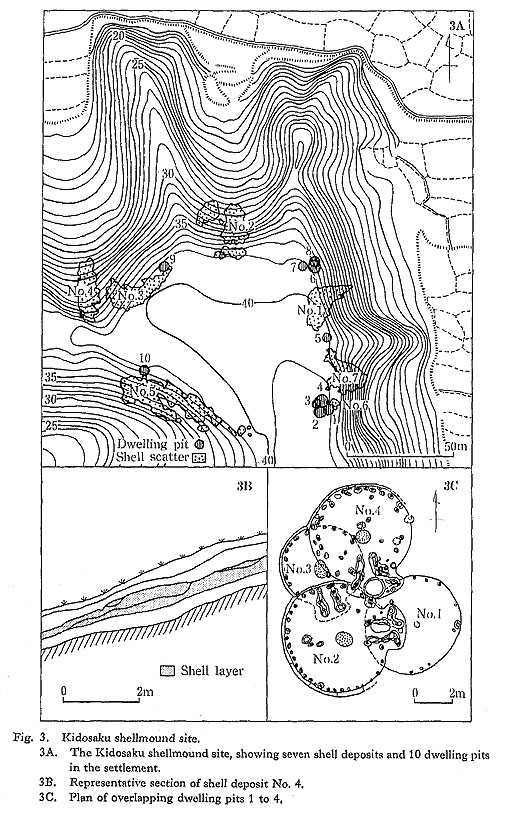
The number of dwellings occupied simultaneously was estimated on the basis of over-lapping between certain structures, and shell deposits found in the fill. Four dwelling pits on the southern edge of the site (dwellings No. 1 to 4) comprise one unit. Successive rebuilding of these dwelling pits appeared to have occurred in one of the two following orders: No. 1→No. 3→No. 2→No. 4, or No. 3→No. l→No. 2→No. 4. Three dwelling pits at the eastern edge of the site (dwellings No. 6 to 8) comprise another unit. Dwelling pits No. 6 and No. 8 overlapped, and dwelling No. 7 was close to these two pits. Dwellings No. 1, 2, 3, 9, and 10 contained shell deposits in their fill, suggesting that they were not
occupied during the latest period of site use. The isolated dwellings, Nos. 5, 9, and 10 seem to have been used independently.
These data indicate that three dwellings, on the average, were occupied contemporaneously. The maximum number of dwellings that might have been occupied contempora-ncously was estimated to be five (Fig. 3D).
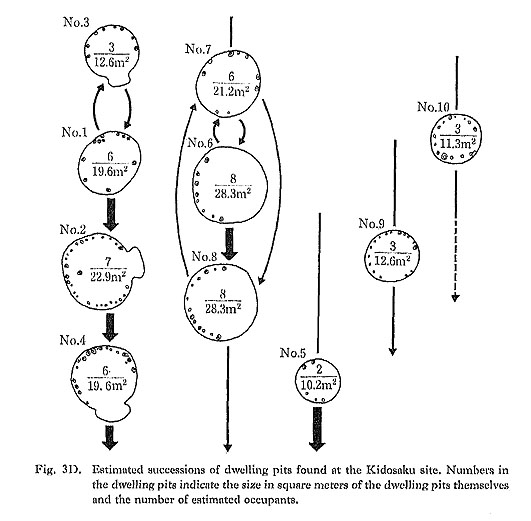
The number of occupants per dwelling has been estimated from the size of the dwelling pits using Sekino's formula (Sekino, 1939):
D=(S-3)/3,
where the number of occupants (D) increases with size of a dwelling pit in square meters (S). The dwelling pits of the Kidosaku site ranged from 11 to 28 m2 in size, hence the number of occupants was estimated at three to nine persons per house (see Fig. 3D). Consequently, the population of this site seems to have been 23 persons at maximum, when five larger dwellings were occupied. It is most probable that the population was roughly 12 persons at the early period of the settlement, later increasing to around 18 persons.
Human remains were found in the shell deposits (Suzuki, 1979). The minimum number of individuals represented by human bones was estimated at nine, although only two formal burials were excavated from shell layers in the fill of abandoned dwelling pits.
Archaeossoological Data
The Kidosaku shellmound site was composed of seven concentrations of shells (shell deposit Nos. 1 to 7) on the fringe of a tongue-shaped plateau or terrace that measured 70 m by 100 m across (see Fig. 3A). These shell deposits spilled down a steep slope of about 20 to 30 degrees.
The area and volume of each shell deposit were calculated by Goda (1979). The concen-trated shell deposits each amounted to 200-400 m2 except for shell deposit No. 6 (Table 1). The maximum thickness of any shell layer was about 1 m, and thickness usually fell within a range of 20 to 40 cm. Hence, the volume of each shell deposit was calculated at 50 to 100 m3, and the total volume of all seven shell deposits is estimated at 450 m3.
Molluscan Remains. ——The molluscan assemblage was analyzed by Komiya (1979) using block samples taken from the corner of each 2 m × 2 m excavation unit in the grid covering each shell deposit. The size of a sample block was 50 cm by 50 cm square, and it was usually 5 cm thick. The molluscan remains identified in the block samples comprised 22 species, and all species were members of a typical tidal assemblage from a sandy bottom.
Among the molluscan remains, the tiny gastropod Umbonium monoliferum was dominant in number of individuals. The clam Meretrix lusoria was dominant among bivalves and is thought to represent the most important molluscan food resource of the site. A total of 5,920,000 valves representing 3,000,000 individual clams were reported from the site. Shell heights of the clams in this site were 25 to 35 mm, indicating a wet weight of living clams of 10 to 15 g each. Hence, the total weight of clams found in the site is estimated at roughly 30 to 45 tons.
Fish Remains. —Small fragments of animal bones, especially fish remains, were detected in block samples which were processed by wet-screening using 1 mm, 2 mm, and 4 mm sieves. Fish remains were identified by Komiya (1979). The assemblage of fish remains consisted mainly of embayment species, such as Cluperioidei sp., Trachurus sp., Hemiraphus sp., and Acanthopagrus sp. Fish remains identified were usually fragments of vertebra, making it difficult to estimate the minimum number of individuals. Therefore, only the number of premaxillary bones of Acanthopagrus sp.(probably A. schlegeli) is listed in Table 1.
Mammalian Remains. —Other animal remains found in the shell deposits were collected individually, their locations three-dimensionally plotted and recorded. Mammalian, avian, and reptile remains were identified by Suwa et al. (1979). The mammalian assem-blage, consisted mainly of species of the warm temperate zone: wild boar (Sus scrofa leuco-myotack) and sika deer (Cervus nippon) were both dominant in this site, as in other Jomon sites in central Japan. The minimum number of individual sika deer was 21 individuals accounted for by lower M2 teeth, or 29 individuals counted at the distal humerus. The total number of individual deer found among seven deposits was 38 (see Table 1).
The wild boar was represented by 36 individuals in total. Besides these larger animals, wild rabbit (Lepus brachyurus) and raccoon dog (Nyctereutus procyonoides) were also commonly found.
Complex cusp patterns on molar teeth and their wear conditions are useful for pairing separated mandibles of an individual deer or wild boar. Applying this pairing technique to the deer mandibles found in the Kidosaku site, only one matching pair of mandibles was identified among the 16 left and 11 right mandibles analyzed. The original population can be estimated using the probability formula (Krantz, 1968; Koike, 1979b):
N=R×L/4p,
where the number of individuals (N) is estimated based on the number of right mimdibles (R) and left mandibles (L) analyzed and the number of paired mandibles (p). Using this formula, the deer population was estimated at about 100 individuals, considerably higher than the preserved population counted by the minimum number method. Poor preservation of bone is assumed to be due partly to the acidic humus soil of the site, and partly to domesticated dogs kept in the settlement.
Reconstruction of Marine and Terrestrial Environments
Paleotemperature Change
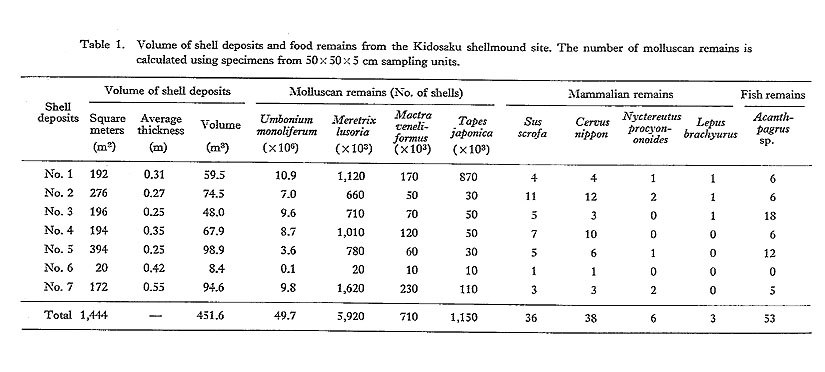
An attempt was made to detect secular changes in water temperature during the Holocene, based on the oxygen isotope ratios of bivalve shells, Meretrix lamarckii Deshayes (Chinzei et al., 1981). The oceanic clam, which is limited to well-sorted sandy bottoms of around 5 m depth, is commonly found in the shell-middens and natural shell beds along Miura and Kujukuri coasts in the South Kanto region.
An example of the oxygen isotope measurements and growth rate analyses is shown in Fig. 4B. The oxygen isotope profile of the shell specimen accords well with the growth rate patterns as established by direct counting of growth-lines; the minimum peaks of the isotope values, i.e., the maxima of apparent water temperature, correspond with the maxima of growth rates. The correspondence strongly suggests that the oxygen isotope variations primarily reflect annual changes of water temperature.
For the oxygen isotope analysis of shells from the Miura and Kujukuri regions, over 800 samples were taken from 21 shell specimens, using a diamond drill to accurately remove shell material from specific growth lines. Among the specimens analyzed, 13 were obtained from shell middens, seven were from natural shell beds, and one was a Recent living specimen.
A clear tendency toward secular change in isotope ratios is seen in the results from the South Kanto area (Fig. 4A). The oldest specimen, from about 8000 B.P., shows the highest 18O concentration, or the lowest isotopic temperature, among the specimens analyzed. The specimen dated ca. 7000 B.P. exhibits the lowest value. The isotope values increase successively from 7000 B.P. to about 4500 B.P., followed by a slight decrease after 4000 B.P. A small low peak may be recognized at about 2000 B.P.
The lowest apparent temperature at 9000 B.P. represents the early phase of postglacial climatic warming, and the peak at 7000 B.P. corresponds to the high water temperature of the Hypsithermal stage. Decrease in sea water temperature about 4500 B.P. is also recog-nized on the basis of shallow marine molluscan fauna in the Holocene sediments; at this time, subtropical species disappeared from central Japan (Matsushima, 1984).
Reconstruction of the Prehistoric Shoreline in the Murata Area
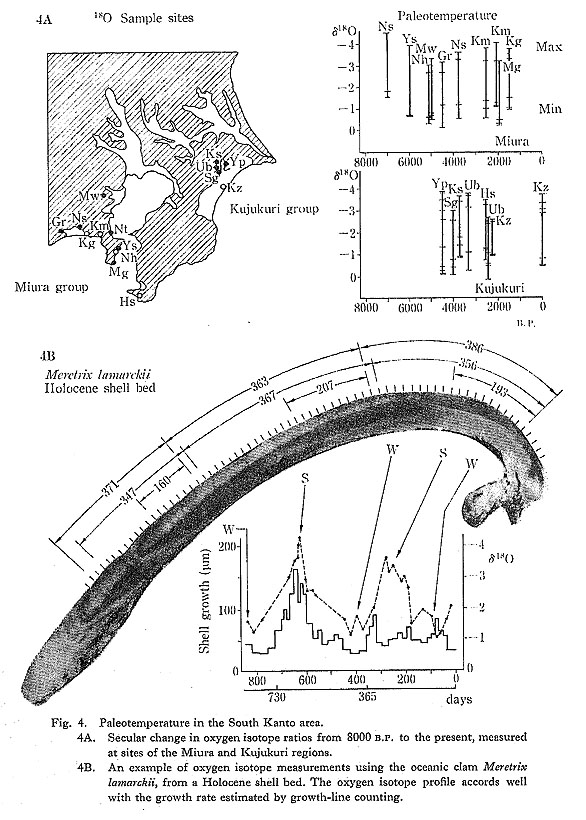
The Holocene sediments in this area were studied by Matsushima (1980) using over 160 core samples distributed from the head to the mouth of the branch valleys of the Murata River system (Fig. 5A, B). the age of the sediments was estimated based on 14C dating of 12 samples. A shell sample obtained from the uppermost part of the marine bed was dated at 7590±860 B.P. (Gak-8016). Although the dates of the peaty humus covering the marine sediments spanned a wide range, they concentrated mostly within 1040 from 5060±160 B.P. (Gak-8020) to 4020±140 B.P. (Gak-8021). Consequently, the period during which the marine sediments were formed is estimated to be 7000-6000 B.P.,and it appears that the area analyzed was covered with peaty humus between 5000 and 4000 B.P.

Based on these 14C dates, the topography of this area developed in three phases: the maximum phase of the marine transgression (about 6500 B.P.), the early post-maximum phase (about 5000 B.P.), and the late post-maximum phase (3500 B.P.).
Maximum Phase of the Jomon Transgression (6500 B.P.)
The paleogeography of ca. 6500 B.P. (Fig. 6A) indicates that the sea had invaded the valleys, forming a complicated shoreline surrounding dissected hills. Sediments accu-mulating in the valleys during this phase were mainly muddy facies of the innermost bay. The upper limit of the sediments containing shells in this area was +2.5 to +3.5 m above the present sea level. Molluscan fossils found at the upper limit consisted mainly of shells which live in the intertidal zone; that is, the -2 to 0 m zone. Therefore the sea level of the maximum phase of the Jomon Transgression in this area can be estimated at +3.5 m to +5.5 m above the present sea level.
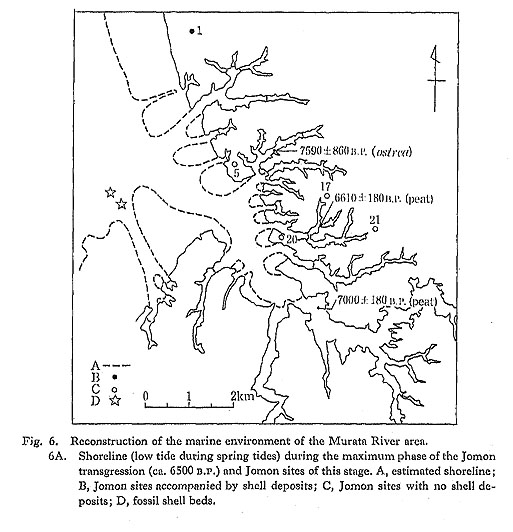
The maximum phase of the Jomon transgression in this area corresponds to the Kayama pottery stage of the late part of the Initial Jomon period. Only four sites contained artifacts
of this stage, and only at the Ohkita shellmound site were they accompanied by shell deposits.
Early Post-maximum phase of the Jomon Transgression (5000 B.P.)
After the maximum phase, the complicated shoreline became simpler and smoother, mainly due to the regression of the sea (Fig. 6B). Reclamation of drowned valleys by fluvial deposits had started even during the early post-maximum phase.
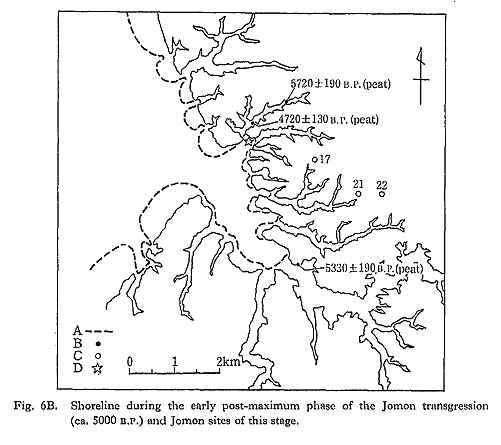
This phase corresponds to the later part of the Early Jomon period (the Kurohama and Moroiso stages). Only three sites are identified as belonging to this stage and none of them is accompanied by shell deposits.
Later Post-maximum Phase of the Jomon Transgression (3500 B.P.)
During the later post-maximum phase, peaty humus deposits appeared in the heads of the valleys, gradually spreading downstream (Fig. 6C). Formation of the upstream peaty deposits indicates that the estuaries had changed into brackish swamps. Regression of the sea prompted the reclamation of estuaries by sandy deposits. The sandy sediments provided a new environment at the mouth of the valley. Molluscan fossils of the intertidal assemblage from sandy bottoms such as Meretrix lusoria, Mactra veneriformis, and Umbonium moniliferum were found at the junction of the Izumiyatsu branch valley and even 3 km upstream from the mouth of the main branch valley.
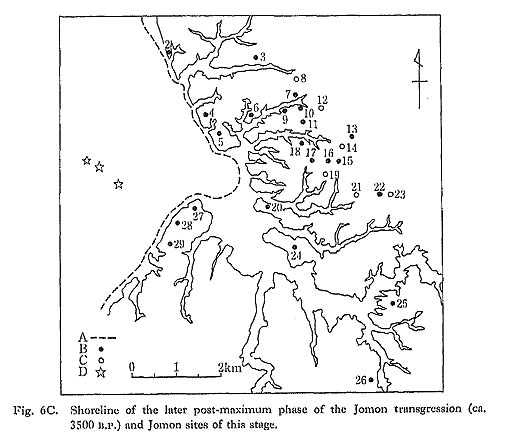
This phase of 3500 B.P. corresponds to the Late Jomon period (Horinouchi and
Kasori B pottery stages). Shellmounds were abundant during this period, and 19 of 23 sites of these stages were accompanied by molluscan remains.
Reconstruction of Prehistoric Vegetation in the Murata River Area
Vegetation analysis was carried out by Tsuji and Minaki (Tsuji et al., 1983) in a branch valley near the Kidosaku shellmound site to reconstruct the flora surrounding the shellmound sites, and also at the conjunction of the branch valleys to reconstruct the regional flora. The whole-core sampling method was used for the analysis, and all the plant remains and volcanic ashes in the peaty sediments were analyzed. Besides the pollen analysis, plant macrofossils were examined to identify more exactly the species found in the pollen assemblage, and to detect edible nuts and cultivated grains. The age of each stratum was determined by both 14C dating and tephrochronology, using ashes found in the sediment core.
The vegetation of this area during the late Holocene after ca. 4500 B.P. consisted of four forest stages (Fig. 7). The first stage, between ca. 4500 and 3500 B.P., is characterized by plentiful deciduous broadleaf trees including Japanese nutmeg (Torreya micifera). The second stage, ca. 3500 to 1900 B.P., shows the maximum development of the warm-temperate evergreen forest. Although the first everygreen trees had appeared by 6000 B.P. at the southern end of the Boso Peninsula, they spread very slowly toward the north, with the result that the major Holocene expansion of the evergreen oak began here about 3000 years later than it did in southwestern Japan. The delay in the expansion of evergreen trees in this region seems not to be due to climatic influence but rather to poor soil conditions caused by the tephras so widely distributed in the Kanto Plain.
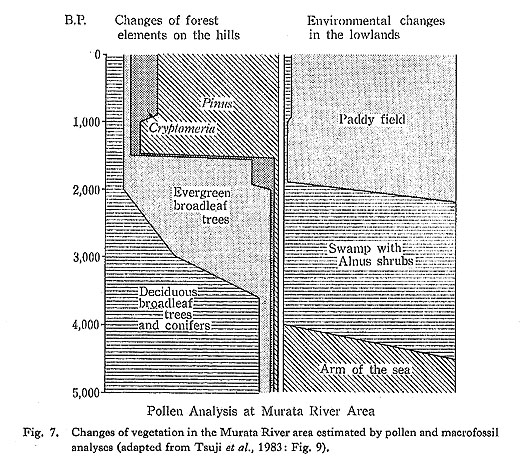
The third stage, ca. 1900 to 1500 B.P., is distinguished by a small expansion of Cryptomeria japonica, which was widely distributed in central Japan by about 2000 B.P. The last stage, ca. 1500 B.P. to present, is characterized by secondary pine forest.
Two phases of vegetational destruction by human activities are recognized in the Murata River area. In the first phase, the marsh alder forest of the branch valleys was destroyed; this was followed by the introduction of rice agriculture. Rice crops (Oryza sativa) were actually detected in a peaty layer about 2 m beneath the surface, which was dated at 2160+140 B.P. (Gak-10306). Weeds and forbs of types commonly accompanying rice were found in the strata, providing useful information on the spread of rice agriculture. The second phase of vegetational destruction saw the disappearance of the natural evergreen oak forest, followed by a rapid expansion of secondary pine.
These data indicate that flora surrounding the shellmound sites during the early stage of the Late Jomon period (4000-3500 B.P.) were composed of plentiful deciduous broadleaf trees in the hills and marsh alder trees in the branch valley lowlands. During the later stage of the Late Jomon period, were the flora gradually changed to an evergreen broadleaf forest a thick, but there still seems to have been a thick forest of deciduous broadleaf trees and conifers on the hilly terraces. Since plant fossils of the secondary forest were rare, it may be that vegetational destruction during the Late Jomon period was restricted to a small range surrounding the settlement.
Plant opal analysis was done by Fujiwara (1984) at the Saihiro shellmound site, which is situated along the next river south from the Murata River. Plant opal remains found in the soil at that site were mainly of sasa (a low-growing bamboo, or bamboo-grass) and other weed-type plants, suggesting that the floor of the forest was rich in sasa, and that the settlement area was quickly covered by sasa during periods of abandonment.
Paleobiomass Analysis of Shellfish Resources
Shellfish Resources in Recent Tidal Flats
The Midori River area (Fig. 6D), which is situated on the western coast of Kyushu facing the Ariake Sea, is rich in molluscs, It is especially famous for the prolific natural reproduction of the clam, Meretrix lusoria. Tidal range in this area is +5.2 to 0.25 m from the tidal datum level during large tides in the lunar cycle. At ebb tides, a large tidal flat extends seaward up to 3.5 km from the mouth of the Midori River.
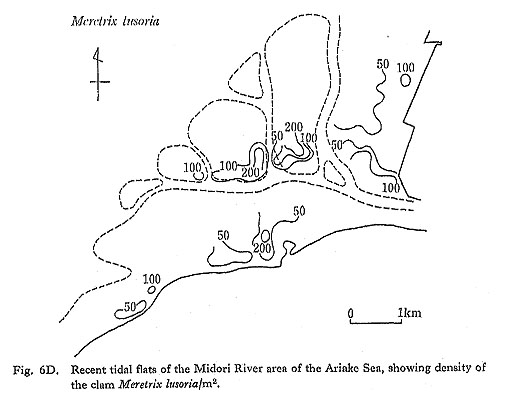
An ecological survey of shellfish resources in the tidal flats of the Ariake Sea was carried out by the Fishery Station of Kumamoto Prefecture (1951). All the shells found within sampling units of 1 m by 1 m at the corners of 200 m grid squares covering a major mollusc field were collected.
Shells of Meretrix lusoria are distributed mainly along the water routes, especially in the off-lying flats. Maximum density of the clam was 700 individuals/m2, and the total amount of the clam resource was calculated roughly at 4500 tons for the 9,000,000 m2 in the area studied (Fig. 6D).
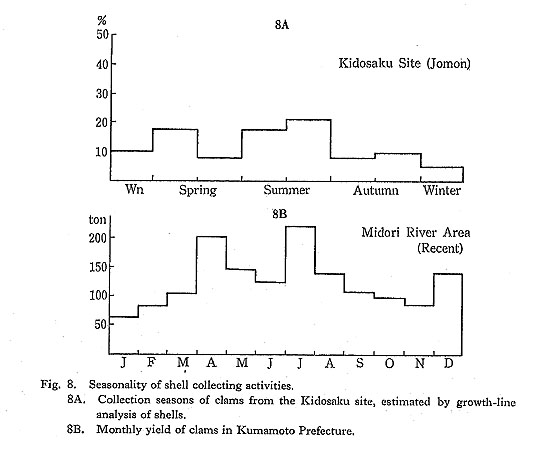
Recent shell collecting activity in the Midori River area is most intensive. More than 200 persons registered with the fishermen's union have been working in this area. Totals for the monthly yield of clams in Kumamoto Prefecture (Fig. 8) show that shell collecting activities continue through the year. Shell collection is especially intensive in spring and early summer, due to the larger and longer exposure of the tidal flats during these seasons.
Shell collecting activities in the Midori River area are highly productive, and an. annual yield of 1,000 to 1,300 tons of clams was attained during the period from 1950 to 1961 (Fishery Station of Kumamoto Prefecture, 1967).
Simulation of Paleobiomass of Shellfish Resources of the Murata River Area
Reconstruction of the prehistoric shoreline in the Murata River area showed that sandy tidal flats developed there during the later post-maximum phase of the Jomon Transgression. Large-scale tidal flats which could have produced great amounts of edible molluscs emerged in an area extending 3 km or more from the conjunction of the branch valleys at the river mouth. Shellmounds in this area were formed in concert with the development of the tidal flats. Although it would be difficult to estimate accurately the area of the Murata River tidal flat at that time, the low neap tidal line is believed to correspond to an old sandy bank formed during the Kofun age (Tsuji et al., 1983: Fig. 8).
The topography during this stage closely resembled that of the Midori River area in the Ariake Sea. The survey of molluscan resources in Kumamoto Prefecture previously referred to showed that about 4,500 tons of living clams were to be found there in an area of 9,000,000 m2. Based on these data, the shellfish resource in the Murata River area during the Late Jomon period can be simulated, as discussed further below.
Collecting Pressure on Shellfish during the Jomon Period
The age composition of the living clam population collected monthly from the Midori River area (Koike, 1980: Fig. V-1) is reflected in Fig. 9, which shows that the specimens studied were composed mainly of 3rd-year-old shells. When shell growth was examined by identifying winter growth minima (Koike, 1982), the first growth minimum appeared at about 9±3 mm in shell height, and the second and third growth minima appeared at 25±5 mm and 31±4 mm in shell height, respectively.
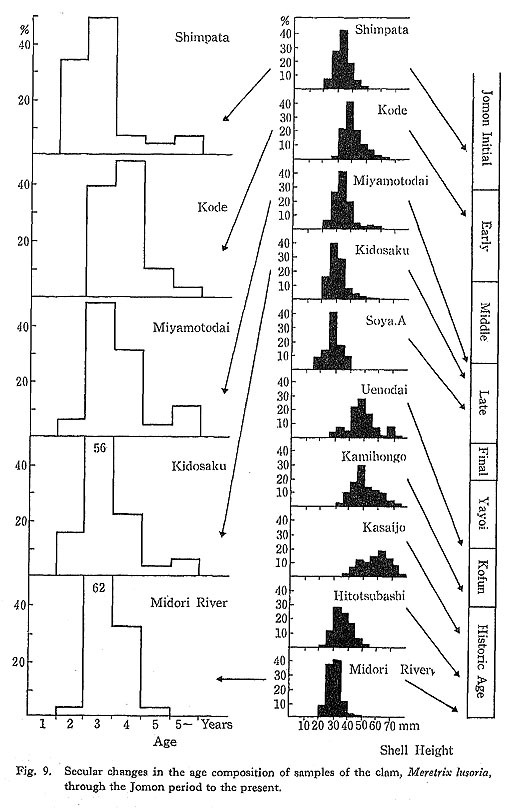
Analyses of age compositions of clams collected from sites dating throughout the Jomon period to the present (Fig. 9) indicate that shells from the Late Jomon period were mostly 3rd-year-old, which is quite similar to the age of recent clams harvested from the Ariake Sea. On the other hand, clams from samples of the Kofun Age were mainly composed of 5th-year-old or older shells. During the subsequent historic age, shellfish were dealt with as a commercial product, and their age composition became as young as those of recent times. These figures make it clear that clams collected during the Late Jomon period were subject to about the same collecting pressure as observed in the Midori River area at the present time.
Discussion
Modern data show that shellfish resources in the Midori River area yield 4,500 tons of clams per year. Based on the fact that the prehistoric habitat for clams in the Murata River area was probably quite comparable to the modern habitat of the Midori River area, and on age composition data which show that prehistoric Murata River area clams were subjected to a collecting pressure as strong as that documented for the modern Midori River area, the shellfish harvest in the Murata River area during the Late Jomon. period is estimated at about 1,000 tons of clams a year, as estimated in the Midori River. In other words, the occupants of any one settlement among the 19 known shellmound sites in the area probably could collect an average of about 50 tons of clams per year.
The Kidoaaku ahellmound site was accompanied by shell deposits totalling 450 m3 in volume, and the number of clams represented was calculated at 3,000,000 individuals, or roughly 30 to 45 tons in total wet weight. The time span of the settlement seems to be very short, within the period of a single pottery stage. Four dwelling pits overlapped with each other, suggesting that the settlement was occupied over a period during which dwellings were rebuilt four times. Based on these data, the time span of occupation at the Kidosaku site is inferred to have been roughly 20 to 30 years. If the average annual consumption of clams is calculated from these data, it comes to about one to two tons per year.
Age composition of clams collected at the site was similar to that for recent times, suggesting that the collection pressure on shellfish was fairly high at the Kidosaku site. Seasonality of clam gathering also shows a similar pattern to that known for modern times (Koike, 1979a and see also Fig. 8), indicating that shellfish-collecting activities at the Kidosaku site continued throughout the year, as intensively as those of modern commercial collectors in the Ariake Sea region.
There is a large gap between one to two tons per year as the actual amount of molluscan remains detected at the Kidosaku site and 50 tons per year as the estimated harvest based on paleobiomass analysis. Although environmental reconstructions using geological and biological data still have difficulties in estimating paleobiomass of shellfish resources, it seems most probable that preservation of molluscan remains in the shellmound sites was simply poor, resulting in a small archaeological recovery. Recently another possibility was illuminated by the excavation of the Isarago shellmound (Education Committee of Minato Ward, Tokyo, 1982): prehistoric people might have had another disposal site for their shells, at a separate shell-collecting camp (Koike, 1983b).
The edible parts of shellfish amount to about 50% of the total wet weight of the animals, and the caloric yield of clam meat is reported as 440 kcal/kg meat. An annual yield of roughly 50 tons of clam was estimated for any single site in the Murata River area; consequently, the annual yield of shellfish meat corresponds to 50,000 kg × 50% meat weight × 440 kcal, or 11,000,000 kcal. The number of occupants at the Kidosaku site was estimated at 12 to 18 individuals, or 15 individuals on the average, as mentioned previously. Their annual minimum caloric needs can be calculated at 15 individuals × 1,300 kcal × 365 days, or 7,000,000 kcal. By this estimate, shellfish collecting could have provided a surplus of food for the occupants, suggesting that dry meat of the shellfish could have been used for trade between coastal and mountainside people.
Paleobiomass Analysis of Game Animals
Recent Deer Populations
Patterns of distribution of the major mammals, such as sika deer, wild boar, black bear, and Japanese serow (Capricornis sp.) have been investigated by the Japanese Agency of Environment of Environment (1979). The existence of animals in each square km was reported for each species, providing useful data for wildlife managers to detect areas threatened by regional extinctions. Although the ecology of wild boar is still unclear, the sika deer and Japanese scrow provide good models for wildlife management.
Wild deer in Nikko National Park, which has the moat stable wild population in the Kanto region, were intensively investigated by Maruyama (1981). The deer population in Nikko is estimated to be between 400 to 600 individuals. The habitat of the deer population in Nikko is about 20 km2, hence the density of the deer population is estimated at 20 to 30 individuals/km2 there. Deer in the Tanzawa area, which is also in the Kanto region, showed a higher density between 40 and 45 individuals/km2 counted in winter (Maruyama, 1974). In general, counting surveys are usually carried out in winter after leaves have fallen. During winter, however, deer aggregate in the lower altitudes of their habitat, resulting in a higher density than the actual average density. Correcting for this bias, the density of deer under natural conditions is thought to be about 10 to 20 individuals/km2.
Simulation of the Paleobiomass of the Deer Population in the Murata River Area
The previously mentioned vegetation analysis for the Murata River area indicated that there was a well-developed deciduous broadleaf forest on the hilly terraces at the time of
the Jomon occupation. The habitat of the sika deer is usually deciduous broadleaf forest or partly coniferous forest, and the main food of modern sika deer is Sasa nipponica, which is common to such forest. Plant opal analysis using soil sampled from the Saihiro shellmound near Kidosaku indicates that the forest floor near the site was covered with sasa at that time. Thus, the flora of the Murata River area during the shellmound stage was a suitable habitat for the sika deer.
The density of living deer in the region at that time is estimated at 10 to 20 individuals/km2, based on the wildlife management data just reviewed. The number of deer has a tendency to increase with the development of secondary forest, but destruction of vegetation by human activities was restricted to a small range surrounding the human settlements during the shellmound stage. Consequently, the deer population during the shellmound stage was probably around 10 individuals/km2.
Hunting Pressure on Excavated Sika Deer during the Jomon Period
The ages of sika deer excavated from 11 sites were determined both by the observation method and the annulation method (Koike and Ohtaishi, 1985). Age determination by observation and measurement is based on tooth replacement, the wear index of molar teeth,
and the appearance of the cervical lines of the molar teeth. Stepwise discriminant analysis was used to establish a criterion for determining the age of individual excavated mandibles.
Age determinations based on analysis of the annual layer of tooth cement were also carried out for well-preserved specimens older than 3.5 years. The winter increment of cement tissue in the sika deer begins to appear in November and December. Formation of the summer increment starts in May and continues until autumn (Ohtaishi, 1980).
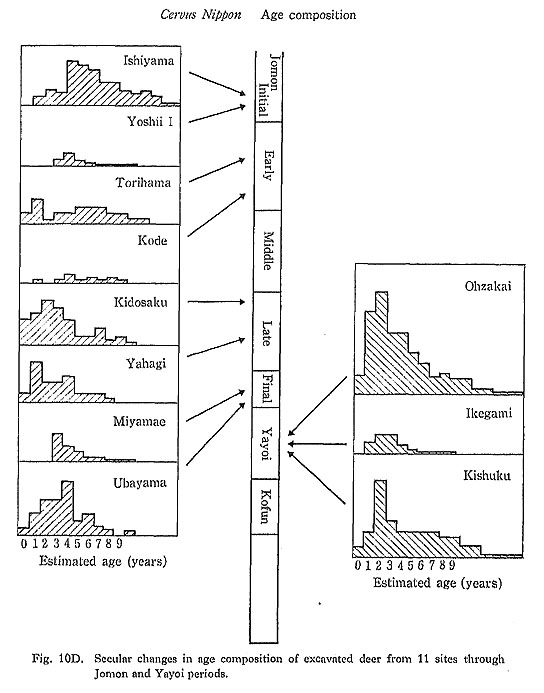
The age composition of excavated deer mandibles from 11 sites (Fig. 10D) changed during the Jomon period. The deer remains from the Initial and Early Jomon periods at the Ishiyama and Torihama shellmound sites were characterized by an abundance of aged individuals. Deer older than five years accounted for 60% of the total analyzed materials. However, in the later periods, data from several sites show that older deer became rare, and deer younger than five years became dominant.
Recent hunted deer killed under natural conditions are in general younger in age composition than deer under protection (Ohtaishi, 1978). Since deer in the Nara Park are fed and well-protected as sacred animals, the age composition of the Nara deer is thought to represent the physiological longevity for the species: the maximum longevity for deer there was 26 years for females and 21 for males (Fig. 10A). Wild deer from Kinkazan Island are protected from hunting but are not fed. Because of these conditions, they are believed to represent the ecological longevity of the species (Fig. 10B): the maximum longevity of the Kinkazan deer was considerably shorter than that of the Nara deer. By contrast, the longevity of hunted wild deer in Hokkaido was only half of the physiological longevity (Fig. 10C). The age composition of the hunted deer showed that the numbers of deer older than five years decreased rapidly, and that deer older than eight years were rare.
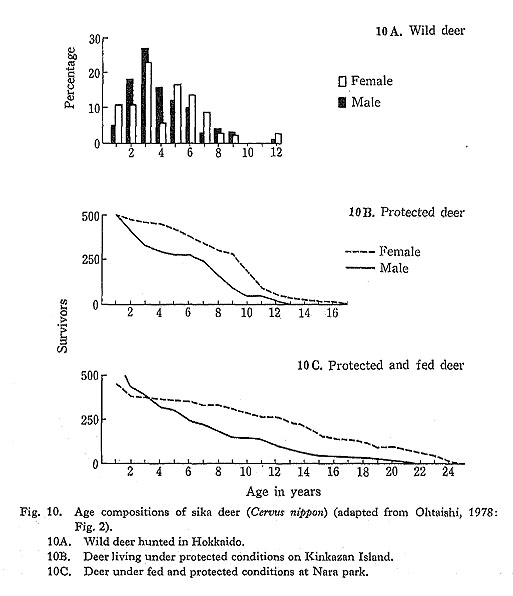
The age composition of the Late Jomon collections showed close similarity with that of recent hunted deer populations in Hokkaido. On the other hand, the age composition of collections from the Initial and Early Jomon periods seems to correspond to that of modern protected deer on Kinkazan Island.
These data suggest that the effect of human hunting activities on the age composition of deer population was negligible during the Initial and Early Jomon Periods, or that during these periods deer were almost free from hunting pressure. Hunting pressure increased during the Late and Final Jomon periods, reaching a level as intense as that on the recent wild populations of deer in Hokkaido, which are hunted to the limit of hunting capacity (see below).
Discussion
In the Kidosaku shellmound site, the minimum number of individual excavated sika deer was 38. When a probability formula using the mandible pairing method was applied to make an alternative estimate, the total number of individuals represented was estimated at about 100. Using the larger estimate, and reckoning the time span of the settlement to be 20 to 30 years, as mentioned above, the occupants' average annual consumption of sika deer as game animals may be calculated at about three to five individuals.
The collection of excavated sika deer from the Kidosaku site was composed of younger animals, showing that fairly high hunting pressure existed, comparable to that experienced by modern wild deer in Hokkaido. The amount of hunting that a wild population can sustain may be estimated based on the age composition using the number of adult females, the pregnancy rate, and, the survival rate of fawns (Clutton-Brock et al., 1982). The estimated hunting capacity for wild deer in Hokkaido seems to be about 10 to 15% of the population (Ohtaishi, 1983).
Based on the analysis that prehistoric hunting pressure at the Kidosaku site was as intensive as that of recent hunting in Hokkaido, where deer are hunted to the limit of hunting capacity at 10 to 15% of the population, and that paleobiomass of the deer population during the shellmound stage seems to have been around 10 individuals per km2, the shellmound people should have obtained deer at the rate of about one to 1.5 individuals/km2.
If the hunting range of the shellmound people in the Murata River area were as same as that of recent hunters in Hokkaido (Ohtaishi, 1983) (one-day-return area on foot), it could be estimated at 10 km×15 km, or 150 km2. Consequently, the potential annual yield of sika deer within that range is estimated at 150-300 animals, or 10-15 animals for each settlement.
Here again there is a gap between the actual number of animal remains counted and the simulation based on environmental reconstruction and the age composition of excavated deer. In the case of large animals, there is an especially strong possibility that butchering often took place at sites distant from the dwelling area. This could account for a smaller-than-expected recovery of animal remains from the excavated site. There is also the problem of limited preservation of bones in the acidic soil.
Average weights of adult deer are 65 kg for males and 45 kg for females (Ohtaishi, 1983). The amount of edible meat is estimated at about 60% of the total weight of the animals, consequently, at about 25 to 30 kg per individual. The total number of calories from deer meat is calculated at 25-30 kg× 10 individuals × 1,300 kcal/kg meat, or roughly 4,000,000 kcal. The annual minimum caloric needs for the Kidosaku site was estimated at 7,000,000 kcal, as mentioned previously. Therefore, the annual caloric intake from sika deer would have been 60% of the annual minimum kcal for the site.
References
- Bourque, B. J. (1975)
- Comments on the late Archaic populations of central Maine; the view from the Turner Farm. Arctic Anthropology, Vol. XII, No. 2, pp. 35-45.
- Bourque, B. J., K. Morris, and A. Spiess (1978)
- Determining the season of death of mammals, teeth from archaeological sites. A new sectioning technique. Science, Vol. 199, pp. 530-531.
- Chinzei, K. et al. (1981)
- Secular changesin oxygen isotope ratios of shells from shell middens with relationship to prehistoric environment. In Studies on Archaeological Sites and Cultural Properties by Means of Natural Sciences, edited by Chairmen group of the Cultural Properties Project. Tokyo: Maruzen. (In Japanese)
- Clutton-Brock, T. H., F. E. Guinness, and S. D. Albon (1982)
- Red Deer. Chicago: The University of Chicago Press.
- Cultural Properties Center of Chiba Prefecture (1979)
- Chiba Newtown Projects, South Area, Report No. 7-Kidosaku Site. Chiba: Cultural Properties Center of Chiba Prefecture. (In Japanese)
- Education Committee of Chiba Prefecture (1983)
- Report on the detailed surveys of the shellmounds in Chiba Prefecture. Chiba: Education Committee of Chiba Prefecture. (In Japanese)
- Education Committee of Minato Ward, Tokyo (1982)
- Isarago Shellmound Site. Tokyo: Education Committee of Minato Ward. (In Japanese)
- Fisheries Experimental Station of Kumamoto Prefecture (1951)
- Reports on the conservation of shellfish resources in tidal flats. Annual Report of Fisheries Experimental Station of Kumamoto Prefecture, 1951. Kumamoto: Kumamoto Prefecture. (In Japanese)
- Fishery Experimental Station of Kumamoto Prefecture (1967)
- Investigation on the amount of damage to the clam Meretrix lusoria and their causes. Annual Report of Fisheries Experimental Stations of Kumamoto Prefecture, 1967. Kumamoto: Kumamoto Prefecture. (In Japanese)
- Fujiwara, H. (1984)
- Personal communication.
- Goda, R. (1979)
- Conclusions. In Chiba Newtown Projects, Southeast Area, Report No. 7-Kido-saku Site (id), pp. 539-542. (In Japanese)
- Hayashi, Y., T. Nishida, and K. Mochizuki (1977)
- Sex and age determination of the Japanese wild boar (Sus scrofa leucomystax) by the lower teeth. The Japanese Journal of Veterinary Science, Vol. 39, pp. 165-174. (In Japanese with English summary)
- Japanese Agency of Environment (1979)
- Report of the Survey on Distribution of Wildlife Animals in Japan. Tokyo: Japanese Environment Agency. (In Japanese)
- Koike, H. (1979a)
- Shell growth rate and collection season of the clams from Kidosaku shell mound. In Chiba Newtown Projects, Southeast Area, Report No. 7-Kidosaku Site (id.), pp. 484-488. (In Japanese)
- Koike, H. (1979b)
- Seasonal dating and valve-pairing technique in shell midden analysis. Journal of Archaeological Science, Vol. 6, No. 1, pp. 63-74.
- Koike, H. (1980)
- Seasonal Dating by Growth-line Counting of the Clam meretrix lusoria. The University Museum, The University of Tokyo, Bulletin 18.
- Koike, H. (1982)
- Growth speed analysis of Meretrix shells, collected in the Hokuriku area along the Japan Sea. Daiyonkikenkyu (The Quaternary Research), Vol. 21-3, pp. 273-282, (In Japanese with English summary)
- Koike, H. (1983a)
- Perspectives in zooarchaeology. The Archaeological Journal (Kokugaku Journal), No. 223, pp. 2-6. (In Japanese)
- Koike, H. (1983b)
- Molluscan analyses. In Studies on the Jomon Culture. Vol. 2, edited by S. Kato et al. Tokyo: Yuzankaku Press, pp. 221-238. (In Japanese)
- Koike, H. and N. Ohtaishi (1985)
- Prehistoric hunting pressure estimated by the age composition. of excavated sika deer (Cervus nippon) using the annual layer of tooth cement. Journal of Archaeological Science, Vol. 12, pp. 443-456.
- Komiya, H. (1979)
- Fish and Molluscan Remains. In Chiba Newtown Projects, Southeast Area, Report No. 7-Kidosaku Site (id.), pp. 372-456. (In Japanese)
- Kranz, G. S. (1968)
- A new method of counting mammal bones. American Journal of Archaeology, Vol. 72, pp. 286-288.
- Maruyama, N. (1974)
- The Japanese deer in Tanzawa mountains. Honyurui kagaku (Mammalian Science), Vol. 28/29, pp. 22-28. (In Japanese)
- Maruyama, N. (1981)
- A study of the seasonal movements and aggregation patterns of sika deer. Bulletin of the Faculty of Agriculture, Tokyo University of Agriculture and Technology, No. 23. (In Japanese with English summary)
- Matsushima, Y. (1980)
- Holocene sediments in Chiharadai Area. In Chiharadai Newtown, Report No. 1, Chiba: Cultural Properties Center of Chiba Prefecture, pp. 94-103. (In Japanese)
- Matsushima, Y. (1984)
- Shallow marine molluscan assemblages of post-glacial period in the Japanese Islands. Bulletin of the Kanagawa Prefectwal Museum, No. 15, pp. 37-109. (In Japanese with English summary)
- Naito, Y. and M. Nishiwaki (1972)
- The growth of two species of the harbour seal in the adjacent waters of Hokkaido. Scientific Report of Whales Research Institute, Vol. 24, pp. 127-144.
- Naito, Y. and S. Konno (1979)
- The post-breeding distributions of ice-breeding harbour seal (Phoca largha) and ribbon seal (Plioca fasciata) in the southern Sea of Okhotsk. Scientific Report of the Whales Research Institute, Vol. 31, pp. 105-119.
- Nishimura, S. (1982)
- Study on the age estimation of the Ryukyu boar (Sus scrofa riukiuanus) from incremental layers in tooth cementum of the first lower molar (1). Journal of Anthropological Society of Nippon, Vol. 90, pp. 143-152. (In Japanese with English summary)
- Ohtaishi, N. (1978)
- Ecological and physiological longevity in mammals. Journal of Mammalian Society of Japan, Vol. 7, pp. 130-134.
- Ohtaishi, N. (1980)
- Determination of sex, age and death-season of recovered remains of sika deer (Cervus nippon) by jaw and tooth cement. Kokogaku to Shizenkagaku (Archaeology and Natural Sciences), Vol. 13, pp. 51-73. (In Japanese with English summary)
- Ohtaishi, N. (1983)
- Deer. In Studies on the Jomon Culture Vol. 2 (id.), pp. 122-135. (In Japanese)
- Ohyi, H. et al. (1980)
- Determination of age, sex and deathsenson of brown bear excavated from the Kabukai A site, Rebun Island, Hokkaido. Bulletin of the Institute for the Study of North Eurasian Cultures, Hokkaido University, Vol. 13, pp. 43-74. (In Japanese with English summary)
- Ohyi, H. et al. (1983)
- Determination of age and mortal-season of northern fur seals excavated from the Kabukai A Site, Rebun Island, Hokkaido. Bulletin of the Institute for the Study of North Eurasian Cultures, Hokkaido University, Vol. 14, pp. 199-240. (In Japanese with English summary)
- Saxon, A. and C. F. W. Higham (1968)
- Identification and interpretation of growth rings in the secondary dental cementum of Ovis aries 1. Nature, Vol. 219, pp. 634-635.
- Sekino, Y. (1939)
- On dwelling pits found from Fukuoka village, Saitama Prefecture and chronology of the dwelling pits. Journal of Anthropological Society of Nippon, Vol. 53, No. 1, pp. 365-382. (In Japanese)
- Spiess, A. (1976)
- Determining season of death of archaeological fauna by analysis of teeth. Arctic, Vol. 29, pp. 53-55.
- Suwa, G. et al. (1979)
- Reptile, avian and mammalian remains from Kidosaku shell mound. In Chiba Newtown Projects, Southeast Area, Report No. 7-Kidosaku Site (id.), pp, 457-467. (In Japanese)
- Suzuki, T. (1979)
- Human remains from Kidosaku shell mound, Loc. 5. In Chiba Newtown Projects, Southest Area, No. 7-Kidosaku Site (id.), pp. 468-469, (In Japanese)
- Taki, J. (1972)
- A tetracycline labelling observation on growth zones in the jaw apparatus of Strongylocentrotus intermedius. Bulletin of the Japanese Society of Scientific Fisheries, Vol. 38, pp. 181-188.l (In Japanese with English summary)
- Tsuji, S. et al. (1983)
- Vegetation and agriculture since the Jomon age with special reference to evidence in the northwest Boso region, central Japan. Daiyonki Kenkyu (The Quaternary Research), Vol. 22, No. 3, pp. 251-266. (In Japanese with English summary)