LATE PLEISTOCENE OSTRACODA FROM THE BOSO PENINSULA, CENTRAL JAPAN
Michiko Yajima
Tokyo Seitoku Gakuen, oji, Tokyo 114
Ostracod assemblage
|
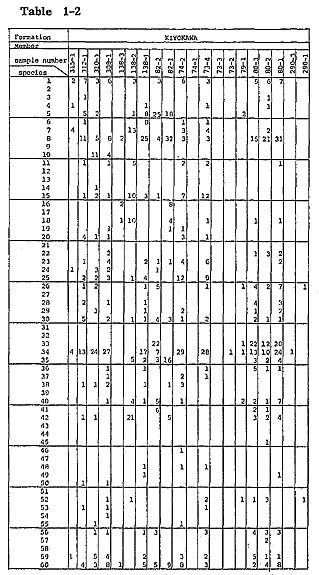
Ostracods representing 121 species were found in 117 samples from 68 localities. A complete list of ostracods from all samples is found in table 1.
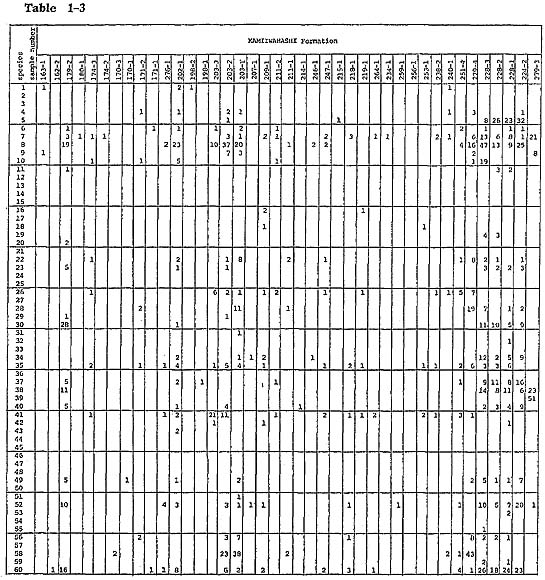
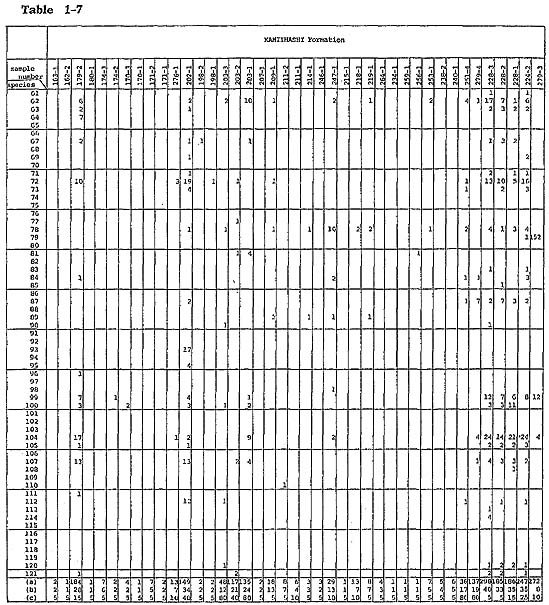
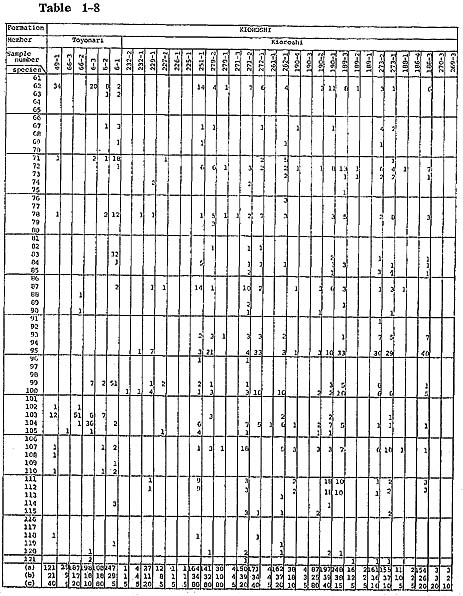
Samples were collected carefully parallel to the bedding plane of fossiliferous sediments to get contemporaneous samples. Burrows filled with subsequent sediments were carefully exiuded. Eighty grams dry weight of sediment from each sample was washed with water through a Tyior's 200-mesh sieve. The washed samples were divided by the quartering method into unit samples which contain approximately 200 ostracods. When the whole washed samples contained far less than 200 ostracods, a washed sample corresponding to only 5 g of sediment was picked because of economy of time. Knowledge on the ecology of modern Japanese species was summarized in the checklist (Hanai et al., 1977). (For additional information, refer to Ikeya and Hanai, 1980 MS.) Unpublished data on the ecology of ostracds of the Aburatsubo Cove were also useful for analytical interpretation of the environment represented by the fossil ostracod assemblages. Yabu Member of Yabu Formation: Ostracods associated with warm-temperate water molluscs, which occur in coarse- to medium-grained sand just below the SY marker-tephra, are characterized by their rareness and high value of Simpson's index of species diversity ( In the coarse-grained sand above the SY marker-tephra, Neonesidea oligodentata, Pontocythere subjaponica, Schizocythere kishinouyei, S. asgao, Loxoconcha optima, Henticytherura cuneata, and Paracytheridea bosoensis occur abundantly. Fifteen sand samples from five localities were analysed. Ostracods are generally abundant. The frequency is approximately 2,800 individuals per 80 g on the average. Three medium-to fine-grained sand samples (samples 305-5, 4, 2) show extreme abundance in the range of 11,000 to 13,000 individuals per 80 g. Species diversity is also very high (λ= 0.062). However, some of abundant species, for example Paracytheridea bosoensis which is associated quite often with Pecten tokyoensis fauna, may be inhabitants of the sand between SY and GoP marker-tephras. A fall in water temperature after the deposition of an SY marker-tephra may be recorded by the presence of a cold-water element, Howeina camptocytheroidea, which occurs in two samples (samples 284-2,1) and another boreal species Finmarchinella (Finmarchinella) uranipponica, which occurs in one sample (sample 305-4). Another sample, 135-3, contains Acanthocythereis? sp. and Robertsonites? reticuliforma, which closely resemble boreal species of Acanthocythereis dunelmensis and Robertsonites tuberculata, respectively. Kamiizumi Member of Yabu Formation: The lower bluish grey silty sand (sample 306-1) with warm sea molluscs living in a depth of 20 to 50 m-for example, Pecten albicans and Chlamys farreri-contains abundant ostracods. Hemicytherura cuneata, Pontocythere subjaponica, and Loxoconcha optima occur abundantly, suggesting that they might be in situ inhabitants or derived from the nearby environment. Munseyella oborozukiyo and Loxoconcha laeta, which are probably warm-water inhabitants, are characteristic. The upper well-sorted, fine-grained sand with warm water molluscs, Umhonium costatum, also contains abundant ostracods. Species diversity being very hgih, the average of three samples (samples 40-2, 1, 138-5) is 0.059. Common sand flat dwellers of the present sea, Schizocythere kishinouyei and Hemicytherura cuneata, occur abundantly. Kiyokawa Formation: Ten sand samples (samples 315-1, 312-1, 310-1, 308-1, 138-3, 2, 1, 82-2, 290-3, 1) were collected from the lower part of the Kiyokawa Formation (lower than TB-7 marker-tephra). Sand just above the basal gravel in the area near Kamiizumi contains an appreciable amount of sand bottom dwellers, Loxoconcha laeta and Hemicytherura cuneata (samples 138-3, 2). The upper horizon of the lowest sand (sample 138-1) contains abundant ostracods with high species diversity (1,000 individuals per 80 g and λ of 0.051). Sand flat dwellers, Pontocythere subjaponica, Hemicytherura cuneata, and Schizocythere kishinouyei, become dominant. In two samples (samples 290-3, 1) from lower black coarse-grained sand near Otabe, ostracods are rarely found. The grain size of the sand is larger than that of ostracod carapaces, and it is possible that their absence resulted from removal by sorting. The fine-grained sand (samples 315-1, 312-1, 310-1, 308-1) just above the silt, which may be correlated with the black clay, has abundant Mactra chinensis lying convex side upward and includes ostracods with very high species diversity near Takinokuchi ( Nine samples (samples 74-2, 1, 73-4, 3, 2, 79-1, 80-3, 2, 1) and one silty sand sample (sample 82-1) were examined from the upper part of this formation. In the silty sand associated with warm-temperate molluscan fauna (sample 82-1) from a bed immediately above TB-7 marker-tephra, ostracods occur commonly (about 900 individuals per 80 g) and species diversity is relatively low (λ=0.122). Loxonconcha optima is dominant. Sand flat dwellers of Hemicytherura cuneata and Pontocythere subjaponica seem less dominant. Aurila kiritsubo and Munseyela oborozukiyo are charac teristic. Five sand samples (samples 74-2, 73-4, 80-3, 2, 1) include abundant ostracods with very high species diversity. They attain from 2,400 to 3,700 individuals per 80 g and In general, no essential difference can be ascertained between ostracod assemblages of sand samples of the Yabu Formation and those of the Kiyokawa Formation. Schizso cythere asagao, Loxoconcha hanachirusato, L. tamakasura, Cytheropteron uchioi, C. miurense, and Xestoleberis sagamiensis decrease, suggesting the dominance of a deeper environment in the Yabu Formation. Loxoconcha kattoi, Aurila hataii, and Callistocythere undulatifacialis increase in their number from the Yabu Formation to the Kiyokawa Formation. Kamiiwahashi Formation: In the lower sand of the Kamiiwahashi Formation, 36 sand samples from 29 localities were obtained. Ostracods are rare in sand samples, ranging from 16 to 300 individuals per 80 g except for sample 179-2 (1,000 individuals per 80 g). Three horizons of molluscan fauna are recognized, and ostracods commonly occur in these three horizons. Pontocythere subjaponica occurs throughout the three horizons. In the lower horizon (samples 203-3, 211-2, 246-1, 247-1, 218-1, 264-1), associated with Panopea japonica in living position, Trachyleberis? tosaensis (sample 203-3) and Hemicytherura cuneata (sample 247-1) occurs characteristically. In the middle horizon (samples 174-3, 170-3, 203-2, 1, 207-1, 209-1, 211-1, 214-1, 215-1, 219-1, 234-1, 259-1, 256-1, 253-1, 238-2, 240-1, 251-4, 279-4) the cold water ostracod Finmarchinella (Finmarchinella) uranipponica is dominant and is associated with alloch thonous molluscan fossils of cold-temperate water inhabitants. Howeina higashimeya ensis, also a cold-water inhabitant, occurs only in this horizon. Callistocythere subjaponica and Aurila cymba are characteristic in the middle horizon. In the upper horizon (samples 163-1, 162-1, 179-2, 180-1, 174-2, 170-1, 171-2, 1, 276-1, 202-1, 198-2, 1), species diversity increases ( In the upper part of the Kamiiwahashi Formation above TAm-5 marker-tephra, brackish water Batillaria-bearing clay (sample 279-3) contains abundant ostracods. Species diversity is very low (λ=0.361). Hemicytherura tricarinata dominates. Brackish shallow water crawlers Spinileberis quadriaculeata and S. fwuyaensis are subordinate yet characteristic. A similar assemblage is observed in the Recent shallow mud bottom of Hamana-ko estuary (Ikeya and Hanai, 1980 MS.). Around Narita, silt (sample 224-2) and sandy silt (samples 228-3, 2, 1) contain abundant ostracods with very high species diversity ( Kioroshi Member of Kioroshi Formation: Among 28 sand samples from 18 localities, 20 sand samples were obtained from the western area of the northern half of the Imbanuma Lake (text-figs. 8, 12). Besides three samples (samples 279-2, 271-3, 272-1) from the basal part with Panopea japonica in living position and 7 samples (samples 271-2, 189-3, 273-2, 1, 186-4, 3) from the lower part with Solen strictus and Tonna luteostoma, 6 samples (samples 279-1, 262-1, 261-1, 190-4, 3, 188-1) from the middle part lower than black scoriaceous sand layer are characterized by dominance of possible Zostera sand dwellers, Loxoconcha hanachirusato and Aurila. cymba, and a sand flat dweller, Pontocythere subjaponica. Species diversity is very high in the basal part (
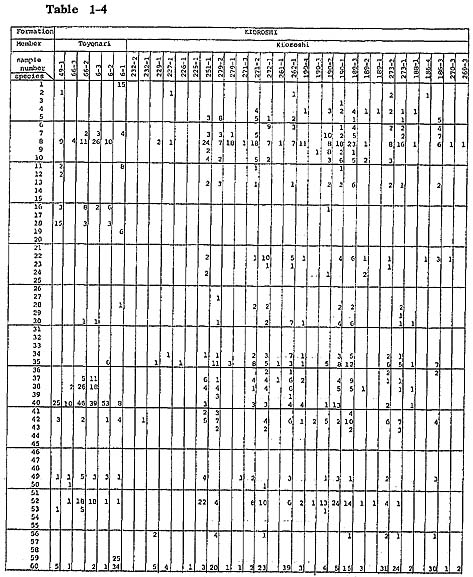
Ostracods from four sand samples (samples 251-1, 190-2, 1, 189-1) from the upper half of the middle part, within and above the black scoriaceous sand layer, are different from those of sand samples lower than the black scoriaceous sand layer in having a relatively abundant mud dweller, Bicornucythere bisanensis, a tide pool dweller, Para doxostoma sohni, and a Zostera sand dweller, Aurila kiritsubo. Species diversity is very high ( Two samples near the type locality of the Kioroshi Formation (samples 270-3, 269-3) and six samples (samples 232-2, 1, 229-1, 227-1, 226-1, 225-1) around Narita, located in the eatern area of the northern half of the Imbanuma Lake, contain ostracods but not in abundance. This is perhaps because they were deposited in a relatively high energy environment. Ostracod assemblage is similar to that of the lower part in the western area of the northern half of Imbanuma Lake. Toyonari Member of Kioroshi Formation: In two samples (samples 66-2, 6-3) from the lower sandy silt with Dosinia angulosa in living position, an inhabitant of the sandy silt bottom, Trachyleberis scabrocuneata, and shallow mud bottom crawlers Cytheromorpha acupunctata, Neomonoceratina microretculate, Spinileberis quadriaculeata, and Bicornucythere bisanensis occur abundantly. Species diversity is relatively low ( In three sand samples (samples 49-1, 6-2, 1) from the upper sand associated with shallow (20 to 50 m deep), warm-temperate molluscan fossils, Zostera sand dwellers, Loxoconcha laeta, Aurila cymba, and A. kiritsubo, and deeper water species, Cornuco quimba tosaensis and Finmarchinella (Barentsovia) japonica, are characteristic. Trachyle beris scabrocuneata is also abundant. Relatively low ( Summary: A cyclic tendency characterizes the upper Pleistocene sedimentation of the surveyed area. Ostracod assemblages vary in accordance with the sequence in the cycle of sedimentation. Most of the samples show high species diversity and do not warrant identification of even dominant autochthonous species or species composition. Yet the following four ostracod assemblages are discernible when one pays special attention to their occurrence in a cyclic sequence. The following four assemblages of ostracods were recognized: subtidal sand, warm water sand, shallow water mud, and brackish water clay. (1) Subtidal sand assemblage: The assemblage is recognizable in the sediments at the maximum transgressive phase. In general, the level subtidal environment was dominant throughout the deposition of the Paleo-Tokyo Bay. The subtidal sand assemblage is found in well-sorted sand containing abundant detached valves of Mactra chinensis. Ostracods are abundant and species diversity is high. This is perhaps due to the addition of allochthonous species from various shallow water environments. The main components of this assemblage are the shallow subtidal sand inhabitants Schizocythere kishinouyei, Hemicytherura cuneata and Pontocythere subjaponica, suggesting the existence of these species in situ or their derivation from nearby environments. This assemblage can be subdivided into two assemblages, based on the abundance of subordinate species, which is perhaps related to water depth. l(a). The deeper water assemblage with which Pecten tokyoensis is associated is characterized by abundant Paracytheridea bosoensis, Cytheropteron miurense, and C. uchioi. Examples are seen in the middle part (high sea level) of the cycle in the middle horizon of the Yabu Member of the Yabu Formation (samples 305-6, 5,4. 3, 2, 1, 284-3, 2, 1, 133-1, 135-4, 3, 2, 1, 120-3, 2, 1). l(b). The shallower water assemblage is characterized by Loxoconcha optima, L. laeta, L. kattoi, Awila cymba, A. kiritsubo, and Callistocythere undulatifacialis. Examples are seen in the lower middle and middle parts of cycles, as exemplified by the Kamiizumi Member of the Yabu Formation (samples 40-2, 1, 138-5, 4), the Kiyokawa Formation (samples 138-3, 2, 1, 290-3, 1, 315-1, 312-1, 310-1, 208-1, 82-2, 74-2,1, 73-4, 3, 2, 79-1, 80-3, 2, 1) and the lower sand of the Kamiiwahashi Formation (samples 163-1, 162-2, 179-2, 180-1, 174-3, 2, 170-3, 1, 171-2, 1, 276-1, 202-1, 198-2, 1, 203-3, 2, 1, 207-1, 209-1, 211-2, 1, 214-1, 246-1, 247-1, 215-1, 218-1, 219-1, 264-1, 234-1, 259-1, 256-1, 253-1, 238-2, 240-1, 251-4, 279-4). (2) Warm water sand assemblage: In the lower formations (Yabu and Kiyokawa Formations) the assemblage is found inserted occasionally in the maximum transgressive sediments with clearly warm-water planktons and molluscs. Therefore, the criterion for classification of this assemblage is different from that of the rest, simply because this assemblage is primarily controlled not by the development of substrata due to eustatic sea level change but by the episodic insertion of warm current inflow at the time of the high sea level. However, in the upper formation (Kioroshi Formation) the assemblage dominates the entire formation. This assemblage is found either in the middle horizon of a cycle corresponding to high sea level phase offshore or nearshore at the base of a cycle, representing the extensively transgressive phase during which the underlying sediments were eroded away. Species diversity is high. The assemblage is subdivisible into s silty sand (20 to 50 m deep) assemblage and a nearshore sand assemblage. 2 (a). A silty sand (20 to 50 m deep) assemblage is found in sand or silty sand associated with warm-temperate water molluscan assemblages. The main components of this assemblage are Loxoconcha laeta, L. optima, Aurila kiritsubo, and deeper water inhabitants of Munseyella oborozukiyo. This assemblage occurs at the middle part (high sea level phase) of a cycle, in the silty sand of the Kamiizumi Member of the Yabu Formation (sample 306-1) and the silty sand of the middle part of the Kiyokawa Formation (sample 82-1). A similar in detail but somewhat modified assemblage characterized by subordinate species, Cornucoquimba tosaensis and Finmarchinella (Barentsovia) japonica, in addition to Munseyella oborozukiyo, is found in the upper sand of the Toyonari Member of the Kioroshi Formation (samples 49-1, 6-2, 1), suggesting a little deeper environment. 2 (b). The shallow nearshore sand assemblage is composed mainly of Loxoconcha hanachirusato, Aurila cymba, and Pontocythere subjaponica. It occurs in the transgressive basal sand of the Kioroshi Member of the Kioroshi Formation associated with Panopea japonica in living position at the base and with Solen strictus and Tonna luteostoma at the lower horizon (samples 279-2, 1, 271-3, 2, 272-1, 189-3, 2, 273-2, 1, 186-4, 3, 262-1, 190-4, 3, 188-1, 270-3, 269-3, 232-2, 1, 229-1, 227-1, 226-1, 225-1). (3) Shallow water mud assemblage: This assemblage is found in the inner bay environment. Its main components are shallow mud bottom crawlers such as Cytheromorpha acupunctata, Neomonoceratina microreticulate, Spinileberis quadriaculeata, and Bicornucythere bisanensis. Associated subordinate species are Trachyleberis scabrocuneata or Paradoxostoma sohni. This assemblage occurs at the horizon just below the high sea-level phase of a cycle of the Toyonari Member of the Kioroshi Formation, in lower sandy silt with Dosinia angulosa in living position (samples 66-3, 2, 6-3). The species diversity is low, suggesting autochthonous occurrence of mud dwellers.The assemblage also occurs in the horizon of a regressive sand of a cycle of Kioroshi Member of Kioroshi Formation in fine-grained sand with Mactra chinensis (samples 251-1, 190-2,1, 189-1). The species diversity is high, suggesting the derivation of mud dwellers from nearby environments. The assemblage is also found in the uppermost horizon of a cycle of Kamiiwahashi Formation, in silt with Dosinia angulosa in living position or sandy silt with Raeta yokohamensis (samples 224-2, 228-3, 2, 1). The relatively high species diversity suggests allochthonous occurrence of sand dwellers. (4) Brackish water clay assemblage: This assemblage occurs in clay with Batillaria zonalis and is exemplified by the uppermost regressive part of a cycle of the Kamiiwahashi Formation (sample 279-3). The species diversity is very low, and ostracod abundance is high. This assemblage is composed of dwellers of brackish water clay of inner bay, represented by Hemicytherura tricarinata, Spinileberis furuyaensis, and S. quadriaculeata. The subtidal sand, warm water sand, and shallow water mud assemblages can be correlated roughly with the sand flat, silty sand, and shallow mud assemblages distinguished by Yajima (1978) in the Kisarazu area. A brackish water clay assemblage is a new addition to the above three assemblages, showing the presence of an area with salinity lower than that of a shallow mud assemblage. Most ostracod species preserved in the sediments of the interglacial period in this area are commonly shallow temperate water inhabitants under the influence of warm water. However, a few characteristic cold-water species, Finmarchinella (Finmarch-nelld) uranipponica, Howeina camptocytheroidea, H. higashimeyaensis, and Robertsonites? reticnliforma may be remnants of the cold-water fauna which may have been dominant in the glacial period, whose sediments are not preserved in this area (Hanai, 1957c; Ishizaki, 1971; Neale, 1974; Okada, 1979). The occurrence of these three species suggests that cold-water influence temporarily remained during the deposition of the Kamiiwahashi Formation and probably of the Yabu and Kiyokawa Formations. |



 where N is the number of specimens, and
where N is the number of specimens, and  is the number of specimens of i-th species). The index attains 0.103 in sample 120-3. The fact suggests that the sample is represented by a mixture of ostracods derived from various environments by postmortem transportation. Dominance of Neonesidea oligodentata and Xestoleberis sagamiensis suggests a warm more-or-less shallow coastal and open sea environment, and the presence of Cytheropteron may suggest a sand bottom more than 30 m deep. However, the abundant occurrence of Pteropoda gives the impression that the coarse-grained fossiliferous sand characterized by lack of fine matrix might have been deposited under the direct influence of warm Kuroshio current flowing into this area from the south.
is the number of specimens of i-th species). The index attains 0.103 in sample 120-3. The fact suggests that the sample is represented by a mixture of ostracods derived from various environments by postmortem transportation. Dominance of Neonesidea oligodentata and Xestoleberis sagamiensis suggests a warm more-or-less shallow coastal and open sea environment, and the presence of Cytheropteron may suggest a sand bottom more than 30 m deep. However, the abundant occurrence of Pteropoda gives the impression that the coarse-grained fossiliferous sand characterized by lack of fine matrix might have been deposited under the direct influence of warm Kuroshio current flowing into this area from the south. =0.065). A sand flat dweller, Schizocythere kishinouyei, dominates, and a cold water inhabitant of Howeina camptocytheroidea occurs in one sample (sample 308-1). Pontocythere subjaponica and Schizocythere asagao are abundant and associated with cold-temperate water molluscs Glycymeris yessoensis in grey medium-grained well-sorted sand (sample 82-2), which is correlated with lower sand distributed around Takinokuchi.
=0.065). A sand flat dweller, Schizocythere kishinouyei, dominates, and a cold water inhabitant of Howeina camptocytheroidea occurs in one sample (sample 308-1). Pontocythere subjaponica and Schizocythere asagao are abundant and associated with cold-temperate water molluscs Glycymeris yessoensis in grey medium-grained well-sorted sand (sample 82-2), which is correlated with lower sand distributed around Takinokuchi.
