FIBULA
Materials
The present materials are eight bones, belonging to five individuals, as follows (Plates 4.53,54,57):
MIr: male, upper one-third is present and some parts of the upper end broken.
MIl: male, shaft only, both ends and a part of the shaft missing.
MIIIr: female, sufficiently complete.
MIIIl: female, lower half only.
MIVr: female, lower three-fourths.
MIVl: female, small fragment from the upper shaft.
MOr: male, shaft only, both ends and the lowest part of the shaft missing.
MPr: male, small fragment from the mid-shaft.



General Character
Since the length of the fibula shows, naturally, the same tendency as in the case of tibia (Tables 4. 31, 32; Fig. 4. 53), there is no need for detailed descriptions except in MO r. The rough estimation for the maximum length in MOr is 320 mm, which is slightly more than that in MIl.
r. The rough estimation for the maximum length in MOr is 320 mm, which is slightly more than that in MIl.



The length-thickness index obtained from the least circumference and the maximum length in MIl, and in MIII r is rather close to the means for the Recent Japanese and the Recent Fukien Formosan and smaller than those for the Neolithic Japanese. The index using the circumference of the mid-shaft is 14.6 in MIl, 14 in MIIIr, 12 in MIVr. The former two approach the means in the Neolithic males and females, respectively, and the latter is close to that in the Recent females. The roughly estimated value of this index in MOr is 19.7, far larger than the means for those Japanese populations.
r is rather close to the means for the Recent Japanese and the Recent Fukien Formosan and smaller than those for the Neolithic Japanese. The index using the circumference of the mid-shaft is 14.6 in MIl, 14 in MIIIr, 12 in MIVr. The former two approach the means in the Neolithic males and females, respectively, and the latter is close to that in the Recent females. The roughly estimated value of this index in MOr is 19.7, far larger than the means for those Japanese populations.
Upper Epiphysis
As far as the available two specimens (MIr, MIIIr) are concerned, there is a wide fovea on the posterior surface of the upper end combined with a well-developed posteromedial ridge for the tibio-fibular ligament. This fovea is presumably for the insertion of the soleus muscle.
Diaphysis
The most striking feature on the side view of the shaft is a pronounced anterior convexity (Fig. 4. 54). It is marked in the anterior border, while the posterior border is nearly straight or slightly concave. In this respect, Minatogawa fibulae bear marked resemblance to those from Fireland cited by Martin (1928). The (anterior) curvature index of the anterior border of the shaft is measured as in tibia, and is 2.3 in MIl, 3.6 in MIIIr and 3.5 in MOr.

The interosseus margin is distinct along the anterior border. This fact indicates that the interosseus membrane is located anteriorly in relation to the bi-articular axis of the fibula (Fig. 4. 54). From an anterior view, the shaft is generally straight with a weak (almost indiscernible) medial flexion at the lower one-third level in MIl and MIIIl.
On the medial surface the area for the tibialis posterior muscle is well defined. It is flat in MIIIrl and MIVrl, while it is concave in MIl and MOr. Interestingly, in these specimens, this area does not expand to reach to the upper end, because its anterior and posterior borders converge at about 2 cm below the upper end. In usual cases, these two borders run upward parallel to each other to the upper epiphysis. In this point also, these specimens resemble the fibula from Fireland mentioned above.
On the lateral surface, the area for the fibularis longus muscle is strongly concave in all the specimens except MIVrl. The cross-section of this region is similar to an L shape in MIIIrl and a  shape in MIl and MOr, implying the strong pressure of the muscles (Fig. 4. 55).
shape in MIl and MOr, implying the strong pressure of the muscles (Fig. 4. 55).

Lower Epiphysis
There is no peculiar character in the lateral malleolar articular surface. The posterior part of the area for the interosseus ligaments is somewhat swollen forming a vertical plane continued from the articular surface. The Fossa malleoli lateralis for the tibiofibular and posterior talo-fibular ligaments is deep in MIIIrl. In MlVr, it is less deep but distinct. The groove for the fibularis muscles is well defined but not deep.
Discussion
Development of the leg muscles
In usual cases, the interosseus space is, from an anterior view, a narrow elongated spindle shape, (Fig. 4. 49). In Minatogawa specimens, however, it is a relatively wide spindle shape whose maximum width is located near the mid-shaft. This condition may somewhat approach that in the great apes.
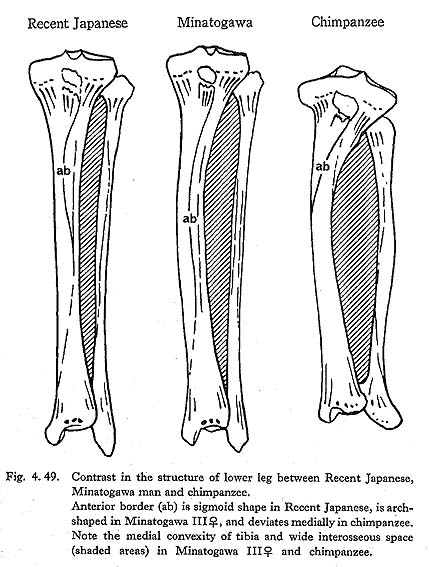
In addition to this fact, there are two characters implying that the leg muscles of Minatogawa man were situated in a distal position, namely, the distal position of the area for the insertion of the tibialis posterior and the shape of the anterior border (Fig. 4. 49). Considering this, the leg muscles which occupy the interosseus space are thought to be well developed, as described before. It is likely that there was a close interaction between these well-developed and distally situated muscles and the anterior convexity of the tibia and fibula.
Distal tibio-fibular joint
In some cases of the Recent Japanese tibiae, the Incisura fibularis is not only deep and narrow but has an inclined surface facing intero-laterally. Therefore, the upward movement of the fibula is perfectly restricted. In this case, on the contrary, this notch is shallow and has a vertical surface, so that the fibula might move upward in the Minatogawa tibio-fibular joint.
TALUS
Materials
There remain only two tali, as follows (Figs. 4. 1, 2; Plates 4. 55, 57):
MIl: male, nearly complete, some parts of the surfaces of the head, neck and the trochlea broken off.
MIIl: female, almost complete, some of the posterior facet for calcaneus missing.



Size
The length of this bone is 47 mm in MIl, 43 mm in MIIl, smaller than the means for the Neolithic and Recent Japanese (Tables 4. 33, 34). The width and height show the same tendency as the length. Accordingly, the Minatogawa tali are slightly smaller in overall size than the Neolithic and Recent Japanese tali.


Head and Neck
The length of head and neck is considerably smaller than those of the Recent and Neolithic Japanese. The navicular facet is somewhat large in proportion to its overall size. It is not as spherical as in Recent Japanese, but it resembles those of the Neolithic Japanese. The ridge bordering this facet to the anterior calcaneal facet is sharp, as is that in the Neolithic Japanese, which shows marked contrast to that in the Recent Japanese. In MIIl, this facet expands posteriorly. In MIl, the expansion does not seem marked, though it is in bad condition.
In both of the present tali, the angle of the head with the trochlear surface is considerably smaller than the means for the Neolithic and Recent Japanese. As for the angle of the neck to the body, that in the present specimens is slightly larger than that in the Neolithic Japanese and somewhat larger than that in the Recent. The subcapitular pit is not visible in these specimens.
Superior Surface
The relative size of the trochlea to overall size is large in MIl as compared with the means of the Neolithic and of the Recent Japanese males. The condition is reversed in MIIl. The trochlear facet is high in both specimens. The sagittal groove of the trochlea is also usual. The lateral and medial margins of the trochlea are at almost the same level when it rests on its basal surface horizontally.
As for MIl, the squatting facets have already been described on the corresponding tibia, but the lateral squatting facet should be described here. However, the preservation of the facets is generally too poor to be able to describe MIl. On the contrary, the description is needed in MIIl, because the preservation is good and the corresponding tibia is not present. The lateral squatting facet is not seen (Fig. 4. 56). A well-developed, smooth-surfaced tuber (Procesus trochlearis, Sewell, 1904) (D') is located on the anterior part of the upper surface of the neck. Usually this tubercle contacts the inferior fossa of the tibia. Accordingly, the tibia of MII presumably had a well developed inferior fossa whose sagittal length was estimated as about 10 mm. The medial squatting facet (B') is poor but distinct. The medial malleolar squatting facet (C') is also more or less developed. Generally, the squatting facets in MIIl are well developed.

The forward prolongation of the trochlear surface along the original curvature is also regarded as an Indication of habitual squatting by many authors (Lisowski et al., 1957; Morimoto, 1959). In MIIl, this prolongation is seen on the medial and lateral part of the trochlea and on the medial malleolar surface, and is fully developed. Such a feature is not clear in MIl because of damage, but from the development of the squatting facets on the corresponding tibia it also may be well developed.
The groove for the flexor hallucis longus is well defined in both specimens. It is located rather laterally and runs obliquely (Fig. 4. 57). The lateral tubercle of posterior process is remarkably developed in MIIl.

Inferior Surface
The anterior and medial facets for calcaneus are united into a single wide and rather flat facet in both MIl and MIIl. The posterior part of this facet in MIIl extends posteriorly to form a small concave facet (Facies inferior accesoria corporis tali, Sewell, 1904) covering the postero-lateral end of the Sulcus tali. The posterior facet for calcaneus is relatively well developed. As in modern Sapiens, its medial (posterior) part is wider than the lateral (anterior). It is not observable because of damage, whether the Facies externa accesoria corporis tali was present or not. Generally, the characteristics of Minatogawa tali bear close resemblance to the Neolithic Japanese tali.
CALCANEUS
Materials
Four calcanei, belonging to three individuals, are preserved (Plates 4. 56, 57):
MIr: male, an upper part of the heel broken off and several pieces missing.
MIl: male, almost complete, some parts of the surface missing.
MIIIr: female, principally complete.
MIVr: female, fairly complete, a part of the body broken off.

Size and Proportion
Compared with the Neolithic and Recent Japanese calcanei, Minatogawa calcanei are very short, more or less narrow and slightly low (Tables 4, 35, 36). Hence the appearance is rather stout. The maximum length in MIl is not only far smaller than the Recent Japanese males but also smaller than the mean for the Neolithic females andequal to that for the Recent Japanese females, though it is male (Fig. 4. 58). The length of the female specimens (MIIIr, MIVr) has the same tendency.



The middle width in MIl falls approximately in between the means for the males and females in both the Neolithic and Recent Japanese. The value for the present female calcanei (MIIr, MIVr) shows the same feature as in the male (MIl), being smaller than those for the Neolithic and Recent Japanese females.
On the contrary, the height in MIrl is slightly less than the mean of the Neolithic males and equivalent to that of the Recent males, as is the case in the female specimens.
Articular Facets
The most impressive feature of the superior view of the Minatogawa calcanei is an exceedingly developed anterior talar facet fused with the middle talar facet (Fig. 4. 59). This fusion is very common in the Neolithic and Recent Japanese (Kiyono and Hirai, 1928b).

In usual cases, the width of the anterior facet is smaller than that of the medial one, but the former is larger than the latter in MIIIr and they are approximately equal in MIl and MIVr. The anterior end of this fused facet overhangs medially and anteriorly on the cuboid facet. This overhanging part continues posteriorly to the Sustentaculum tali. The cuboid facet extends postero-medially under this overhanging portion.
The posterior talar facet is also well developed. In all the specimens the value of the width exceeds the means for the Neolithic and Recent Japanese, though its length is rather small.
Sometimes the facet which contacts with the Facies externa accesoria corporis tali is seen along the anterior border of the posterior facet. It is considered a mark of habitual squatting. All the Minatogawa calcanei have this facet, but not as developed as in the Neolithic Japanese calcanei.
Lateral Surface
The peroneal tubercle is well developed in MIrl and MIIIl. This is not observable in MIVr for breakage. The lateral tubercle is present in all specimens. The connecting bridge between the peroneal tubercle and the lateral tubercle is seen in MIIIr. In MIrl, these two tubercles are joined to form a wide prominent tubercle. The groove for the peroneus longus and brevis is well defined and is divided by the peroneal tubercle.
Other Characters
The medial tubercle is well developed and projects inferiorly and medially in MIl. Its most projected point is located 8 mm below the basal surface. This condition in MIr is not clear because of damage.
In MIVr most of the medial tubercle is dislocated medially and superiorly by a rather complicated fracture (Fig. 4. 60). The posterior end of the medial tubercle is not displaced but the anterior end is displaced about 4 mm. Therefore, this portion is twisted counter-clockwise when the bone is seen from above. Along the fracture line between the medial and lateral tubercles there are several small pieces of compact bones depressed from the original surface. The fracture has been tightly cemented by lime carbonate. This fracture presumably occurred at around the time of death.

Discussion
Subtalar joint
Since the articular facets for the talus are exceedingly developed, it seems that Minatogawa people used the subtalar joints quite well. This means they may have often walked or run on uneven, rough ground. This feature may also be the same in the Neolithic Japanese.
Shortness of the heel
As described above, the Minatogawa calcanei are quite unique for their shortness in total length. This shortness is due mainly to the small size of the heel (Fig. 4. 59); for instance, the length of the heel is 33% of the total length in MIl, 33% in MIIIr and 37% in MIVr. It is well known that some Neanderthals also had short heels, but the ratio of Minatogawa people with this trait exceeds the Neanderthals (40% or slightly more, according to McCown and Keith, 1939). The functional interpretation of this shortness of the heel will be discussed later as "leverage of the foot."
Position and development of the lateral tubercle
Two positions of the lateral tubercle are known.
First: The lateral tubercle is situated high on the lateral side of the calcaneus and its surface is similar to the other lateral surface (Fig. 4. 57). It provides an attachment area for the planter aponeurosis or abductor digiti minimi muscle but it does not support the body weight.
Second: It is situated low on the infero-lateral corner of the heel and its surface bears an exact resemblance to that of the medial tubercle. Usually its posterior part continues to the medial tubercle. It is used not only for the attachment of the aponeurosis or muscles but also for the support of body weight, as is the medial tubercle.
In the first type, the lateral tubercle is usually rather poor while it is commonly well developed in the second type. Considering that this tubercle is not seen in great apes and that it serves as body support when its position is low, the second type seems to be progressive.
In Japanese, the second type is usual but the first is also seen, particularly in females (Kiyono and Hirai, 1928b), In the Minatogawa specimens, the first type is seen in MIrl and MIVr and the second one in MIIIr.
OTHER FOOT BONES
Os naviculare
Only one incomplete left bone from MIII has remained (Plates 4. 28, 55). Its posterior facet is almost complete, though some parts of the tubercle and anterior facet are broken off. The width of the remaining part is 31 mm and the original width was estimated as 33 mm, smaller than the means for Neolithic or Recent Japanese females (Table 4. 37). The height is 21 mm, which is far smaller than that for those people. The minimum thickness is also smaller.


In proportion to the overall size, the posterior facet is also smaller. As is usual, the anterior facets for the second and third cuneiforms are flat or slightly concave in the upper part.
Generally, the character of this navicular has no significant difference from those of the Recent Japanese, but it is quite small.
Os cuboideum
Only one left bone from MI is preserved in nearly complete condition (Plates 4. 28, 55).
The most impressive character of this bone is a steep inclination of the posterior facet; its angle to the anterior facet is only 45°. The length of lateral surface is 13.5 mm, which is only 41% of the lower length of the medial surface (33 mm). This value (41%) is far smaller than the means for the Neolithic and Recent Japanese males (55.6% and 50.6%, respectively). The upper length of the medial surface is 29 mm and its ratio to the length of lateral surface is 47%, which is intermediate between those for the Neanderthals (41.6% in 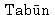 I, 42.8% in
I, 42.8% in  IV) and modern Sapiens (55.5% in Bushman, 50.0% in Sikh, according to McCown and Keith, 1939).
IV) and modern Sapiens (55.5% in Bushman, 50.0% in Sikh, according to McCown and Keith, 1939).
The ratio between the width of the posterior facet and the upper length of medial surface is 90%, showing the same tendency as the ratio just mentioned. The facet for the third cuneiform is distinct but the facet for the navicular is not present.
The calcanean process is marked. The groove for the peroneus longus is deep combined with the pronounced retroperoneal ridge. The height of the ridge from the base of the groove is 10 mm at the lateral end and 5 mm at the medial end. The facet for the sesamoid cartilage is well developed.
Metatarsals
We have nine bones; five belong to MI and three to MIII, and one is isolated (Q) (Plates 4. 28, 58).

First metatarsal
MIr: male, distal three-fourths.
MIIIr: female, nearly complete, some part of the proximal end broken off.
The estimated total length of MIr (56 mm) falls in the middle between the means for the Neolithic and Recent Japanese males (Table. 4. 38). The length of MIIIr (51 mm) is smaller than the mean for the Neolithic Japanese females and close to those of the Recent females and I (Table 4. 39). As for the stoutness, MIr falls between those of Sikh and Bushman, MIIIr is less than that of I.


The area for the insertion of the peroneus longus muscle is well defined forming a sharp pit in MIIIr. The articular surface for the sesamoids of the flexor hallucis brevis and adductor hallucis is somewhat wide extending posteriorly in MIr. In both of the specimens, the cross-section of mid-shaft is triangular as in usual cases of modern Sapiens.
Second metatarsal
MIl: male, distal one-fourth missing.
MIIIr: female, distal end missing.
The roughly estimated values for the total length in MIl and MIIIr are less than the mean for the Neolithic Japanese but not less than those for the Recent Japanese (Tables 4. 38, 39). Both shafts are triangular in cross-section having sharp edges. The shaft of MIl seems to be thick in relation to its length.
The facets for the second cuneiforms are usual in both specimens. The beveled facets for the third cuneiforms are also seen; their width is 3 mm in both MIl and MIIIr.
Third metatarsal
MIr: male, approximately complete.
MIIIr: female, proximal one-third.
The length of MIr (66 mm) is a little less than the mean for the Neolithic Japanese male and larger than that for the Recent ones. The shaft of MIr is extremely thick, though its length is not large. The shape of the cross-section of mid-shaft is flattened transversely. No peculiar character is seen in MIIIr.
Fourth metatarsal
Only one shaft of this bone is preserved, namely, MQl. It was at first regarded as belonging to MI. But from the diameter of mid-shaft and estimated length, this specimen is presumed to belong to a female, although it may not belong to MI. No special character is evident except for its transverse flatness of the shaft.
Fifth metatarsals
MIr: male, complete.
MIl: male, principally complete.
These are very long and stout compared with those of the Japanese. Their total length is longer than the means for the Neolithic and Recent Japanese males. Their diameters are equivalent to those of the stout specimens from Sikh (McCown and Keith, 1939). Both the shafts are quite curved laterally in the distal parts. In both specimens, the articular facet for the cuboid is well developed and extends posteriorly covering most of the tuberosity. This facet of MII fits well with the corresponding facet of the cuboid. The area for the peroneus brevis is well defined on the tuberosity.
Phalanges
We have three proximal phalanges, one from MI and two from MIII (Plates 4. 28, 58).
On the larger bone from MIII, the inclination of the distal end is not present. Its length (26.5 mm) is slightly less than the mean of the second bone of the Neolithic Japanese females (27.1 mm) and larger than the third one (24.3 mm). It seems that this bone is a second proximal phalanx, but it is not known whether it is right or left.
The smaller bone from MIII is short (20.5) and the postero-lateral inclination of the distal facet is marked. It is presumed to be a right fifth proximal phalanx.
The bone from MI is 23 mm in length. This value fits the mean of the fourth bone of the Neolithic Japanese males (23.2 mm). The postero-lateral inclination of the distal facet is, however, very slight and as a male, it is too slender. Accordingly, it might belong to a female second or third proximal phalanx (MR).
Leverage of the foot
As mentioned above, the most striking feature in the Minatogawa feet is the relative shortness of the heel in spite of their rather stout build. We will now discuss the leverage of the foot.
Since there are no data on the length of the heel in the Japanese, we will use the total length of the calcaneus as a representative indication of force arm length. As representative of load arm length, we will use the length of the cuboid and third metatarsal for MI (male), or the thickness of the navicular and length of the second metatarsal for MIII (female). As the length of the cuboid, the mean of the length of lateral surface and lower length of the medial surface is used, and for the thickness of the navicular, the mean of the minimum and maximum thicknesses. The ratio between the representatives of the load and force arms (leverage) is considerably higher in Minatogawa feet than in Japanese (Table 4. 40). These high ratios mean that the movement of the tiptoe is relatively higher in verosity and wider in range than those of the Japanese. Accordingly, if the leg muscles of Minatogawa man were not strong in relation to their body weight, they could run neither fast nor easily on uneven ground. But it seems that the muscles were strong. The relative shortness and stoutness of the heel is also seen in Mt. Carmel man, and McCown and Keith stated that it relates to their ample calf muscles (1939).
