動物学レファレンス標本について
飯島魁教授のガラス海綿類の研究とその後の展開
|
村上彰 浜松医科大学 |
1 はじめに正確には記憶していないが、昭和51(1976)年より平成3(1991)年に至る約16年間、当時の理学部生物学科動物学コースで、高橋景一教授のお手伝いとして、無脊椎動物学の講義を担当させていただいた。現在まで、いく種類かの講義をしてきたが、最も学び・教える喜びを味わうことのできたのは、ためらいもなくこの講義であったと断言できる。研究者としての私の専門は、動物生理学または比較生理学であり、分類学や系統学ではなかったが、形態と機能の関係に注目して無脊椎動物の話をすることによって、授業時問ごとに私自身の目が開かれていく興奮を味わうことができた。このような機会を与えられた幸運を今でも深く感謝している。この講義を始めるに当たって、何冊かの無脊椎動物学の教科書を選んで自分なりに目を通したが、例えばR・M・アレクサンダー(R. McNeill Alexander)やG・A・カーカット(G. A. Kerkut)のように生物物理学(力学)や生理学の大家と思っていた人達が、無脊椎動物学や脊椎動物学の立派な教科書を著したり、分類学に視点をおいた標準的な無脊椎動物学教科書の大規模な改訂をしたりしていることを知り、深い感動を覚えたものである。特に、アレクサンダーの教科書(The Invertebrates, Cambridge Univ. Press 1979)は、講義の参考書としてだけではなくテキストとしても使用させていただき、多大の影響を受けた。機能を中心とした系統学が如何に好奇心を刺激する面白いものであるかを改めて教えられた。そして、その興奮の何割かは、その話題の中心となる実物の標本と対面することによってさらにかき立てられたものと確信している。 動物学教室の講義室には、講義用と思われる標本があり、展示と言うよりはむしろ無造作に保存されているという態で、棚の中に雑然と置かれていた。その中には、由来のはっきりしないものも含まれていたが、原始的な腹足類(巻貝)のオキナエビスガイ(Mikadotrochus beyrichi)など、よく知られた逸話が伝えられている標本も何気なく置かれてあった。また、三崎の油壷のような観光地の土産物屋にも売っているような貝殻と並んで、オウムガイ(Nautilus pompilius)の軟体部の液浸標本もあった。オウムガイは、生きている化石としてよく知られた種で、その浮力調節機構が、実は浸透圧調節の特異な例であることから、動物生理学の講義でも使わせていただいた。さらに同じ標本を、二対の鯨を持った古いタイプの頭足類の生き残りとして、無脊椎動物学の講義でも重ねて使用させていただいたものである。また、トグロコウイカ(Spirula spirula)は、鰓が一対しかない新しいタイプの頭足類であるにもかかわらず、アンモナイトやオウムガイのような平巻きの甲を持っている変わったイカであるが、その小さな甲の標本を見つけた時には、喜びに思わず声を挙げたものである。私は、それらの多彩な標本で大いに楽しませていただいたが、自らその整理を手掛けたこともなく、全容を解説する立場にもない。この稿では、私にとって特に印象の深かったカイロウドウケツ(ガラス海綿類、六方海綿綱 Hexactinellida)の標本にまつわるお話しをお伝えすることに止めたい。 2 カイロウドウケツの標本にあいた窓総合研究博物館には、動物学教室由来の美しいカイロウドウケツ[挿図1]を始めとして、多くの海綿類の標本が保存されている。それらの多くは、長年にわたり海綿類の研究を集中的に行なった飯島魁(Isao Ijima)教授の業績に関わるものである。動物学教室の講義室に、あまり見栄えのしない海綿の標本があり、そのほぼ中央部に組織を矩形に切り取った跡があった[挿図2]。それを最初に見た時は、それが何の痕跡かと気にはしても、その由来を調べてみることもなく時を過ごした。しかし、後に、それは飯島が海綿類の形態を研究するために自ら組織標本を切り取った跡であることを知り、その研究がその後どのように発展していったかを悟った時に受けた深い感銘を今も忘れることができない。飯島は、漁師から買い上げた標本ばかりでなく、三崎臨海実験所を根拠地として自ら底延縄(そこはえなわ)を使って採集を行ない、それらの標本を基に、多くの新種を記載し、特に六方海綿類の分類の基礎を築いた。彼の研究の経過と海綿に関する主な業績(論文)については、研究総合資料館展示解説[堀越1983]に紹介されている。また飯島は、採れた標本を船上で直ちに固定して組織研究用の標本を作成し、光学顕微鏡による詳細な形態学的、組織学的研究も行ない、多くの重要な観察を行なった。
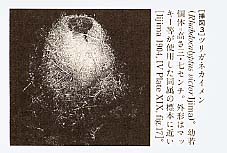
ここに取り上げたいのは、彼の組織学的な観察結果の再確認の話である。飯島は、光学顕微鏡による観察によって、ガラス海綿類の組織が連統した多核体(シンシチウム)で構成されていることを記載していたのである。私の専門は動物生理学で、繊毛運動の制御機構を研究テーマとしていた。私はかねてから、カナダのG・0・マッキー(G. O. Mackie)等の、クラゲやホヤ等の無脊椎動物における神経様伝導(neuroid transmission 類神経伝達)の研究には興味を持ち、注目していた。神経様伝導というのは、神経細胞を介さないで、上皮組織を電位変化が伝導し情報を伝える現象をいう。ある日、高橋景一教授がセミナーでガラス海綿に関するマッキー等の研究を紹介されたが、その内容は驚いたことに、何と神経の網状説(reticular theory)復活の話であったのである。 3 ニューロン説と網状説この話の興味の焦点がどこにあるかを説明するためには、神経系の構成に関する論争の歴史をごく簡単に振り返る必要がある。現在では、神経単位(ニューロン neuron)という語は、一つの神経細胞を意味する単語として広く使われている。しかし、これが定着する以前は、中枢神経系(脳や脊髄)の神経回路が、どのような構成単位によって成立しているかが論争の的であった。ニューロン説(neuron theory)では、神経細胞が個々に独立した活性(興奮状態)を持つとしたが、対立するC・ゴルジ(C. Golgi)等の網状説では、神経細胞は構造として連続しており、神経興奮は境界のない網目状の回路を伝わるとした。この論争は、S・R・カハール(S. R. Cajal)等の詳細な神経組織学的研究によってニューロン説に軍配が上がった。この経過については多くの本に紹介されている[萬年1967等]。飯島が、海綿の形態学に熱心に取り組んでいたのは、まさにこれらの対立する両説に決着がつこうとしていた今世紀の初頭のことである。 この話を続けるには、さらに細胞から細胞へ情報が伝えられる機構についてのもう一つの論争に触れる必要がある。現在では、大部分の細胞間のシナプス伝達は、主として軸索末端からのアセチルコリンのような特定の伝達物質の分泌による、いわゆる化学的伝達(chemical transmission)により行なわれていることは、広く知られている。その対立説であった化学物質によらない電気的な伝達機構(electrical transmission)も、心筋や平滑筋だけではなく、幾つかの神経細胞間についても報告されている。これは、細胞の特別な構造を持つ接触部(ギャップ結合 gap junction)を通過する電流が、隣の細胞の膜電位を直接変化させるもので、典型的には全か無かの原則に従う活動電位を伝播させる。結果としてこれらの細胞は、境界のない連続した細胞の集まりのように振舞う。このような細胞の集まりは機能的多核体といわれる。マッキーは、幾つかの例について、形態的には上皮である細胞が、上に述べた神経様伝導によって活動電位を伝播させることを研究していたのである。 4 海綿類の刺激に対する反応とガラス海綿類の水流停止反応海綿動物門は、多細胞動物の中では最も細胞の分化が進んでいない、基本的には壷のような形をした動物の集まりである。海綿動物には、種々のパターンに従って分布する鞭毛室(flagellated chamber)があり、その内壁に、襟細胞、または有襟鞭毛細胞(choanocyte)と呼ばれる特徴的な鞭毛細胞が並んでいる。この襟細胞には、一本の鞭毛と、それを円筒形に囲んで襟のようになった突起の列があり、襟細胞の存在は海綿動物の顕著な特徴の一つとされ、ガラス海綿にも同様の構造が観察される。これは、原生生物(単細胞藻類)の一種である襟鞭毛虫(choanoflagellate)と共通した他に例のない特殊な構造で、海綿動物がそのような藻類から独自に進化したとする考えも出されている。海綿類は、襟細胞の鞭毛運動によって摂食流を作り、その水流に乗って運ばれてくるプランクトンを襟で捕えて餌とする。体表の小さな入水口(incurrent pores)から入った水は、鞭毛室を通り、中央の水路(canal)に集まって出水口(excurrent vent; oatia)より出ていく。海綿動物は運動性に乏しく、入水口や出水口の口径が変化したり、部分的に変形したり、例外的に個体全体が変形するものはあるが、分化した筋細胞や神経細胞はなく、一般的には、協調した運動をするために必要な情報伝達機構はないと考えられている。特にガラス海綿類には、収縮運動とそれを制御する調節機構はないと言われていた。しかし、それを確かめようとしても、この海綿の仲問は通常は深海産であるために、海底を引きずって集めるドレッジや底延縄のような手段でしか採集できず、取れた標本の傷みがひどく、なかなか健全な標本を採集することはできなかった。特に、生理学的な実験を するために必要な標本の入手は極めて難しく、その機能の研究はなかなか進展しなかった。 マッキーらは、ガラス海綿類のツリガネカイメン(Rhabdocalyptus victor Ijima,1897、幼若個体)[挿図3]の近縁種、Rhabdocalyptus dawsoni (Lambe 1873)が、カナダのブリティッシュコロンビアの沿岸では20—30メートルと比較的浅い海域に生息することに注目し、スキューバ潜水によってほぼ完全な標本を採集することができた。この海綿は極めて壊れやすいけれども、注意深く扱い、低温(摂氏15度以下)に保った新鮮な海水中では数週間にわたって飼育可能であった。彼らはこの標本を利用して生理学的実験を行ない、ガラス海綿類には、海水を流したり止めたりする調節機構があることを示したのである[Lawn et al. 1981]。使用した標本の外形は、挿図3に示した種の幼若個体に近い。
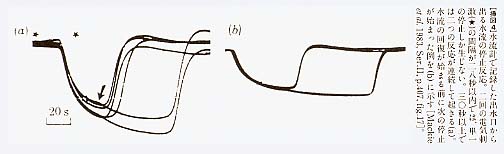
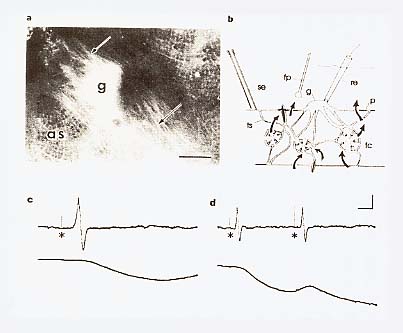
マッキーらは、上のようにして得たガラス海綿の出水口に水流計を置き、出てくる水流の強さを計った。水流計は、サーミスターを電流で暖め、そこから水流によって熱が奪われるために低下する温度を測定するという原理を用いた、感度の良いものである。彼らは、これで測定された水流が自発的に停止することがあるばかりでなく、海綿を入れたタンクを叩いたり海綿をそっとつつく等の機械的な刺激や、電流刺激を与えても停止することを発見した[Lawn et al. 1981]。マッキーらは、以下に述べるような実験から、この水流の停止が、ガラス海綿全体の鞭毛が一斉に止ることによると結論したのである。反応は刺激後20秒から50秒で現れ、水流の停止は30秒から100秒間持続した。時には不完全な水流の停止がみられたが、反応はほぼ「全か無かの原則」に従うものであった。繰り返し刺激によって反応の持続時間は長引いたが、反応が現れるまでの時間(潜時)は変化しなかった。また、海綿のどの部分を刺激しても一定の反応が生じた。この結果は、この海綿に刺激伝導系が存在することを示唆するものである。彼らは、出水口周辺の体壁の一部を、一端を本体に付けたまま短冊型に切って引き出した標本を作り、出水口からの水流が停止するまでの反応時間が、刺激点を移動したときにどのくらい変化するかを測定し、その差から刺激伝導速度を計算したところ、ほぼ毎秒二ミリメートルの値を得た。水温は摂氏11度であった。その後、マッキーらは、さらに感度の良い水流計によって、海綿の体壁を矩形に切り出した標本から流出する水流を直接計測し、水流が停止する反応のより詳細な性質を明らかにすることができた[Mackie et al. 1983]。まず、体壁から流出する水流が停止する波形を記録し、水流は刺激に続いて全部位でほぼ一斉に完全停止し、回復時には、水流は30秒程度にわたって次第に増加してくることを示した。反応を繰り返すと疲労が現われ、水流の停止時間は長くなり、回復する水流の立ち上がり速度は遅くなる。刺激部位を水流の測定場所に近付けることによって、反応時間は1から2秒まで短くなる。水流計の時定数は約1秒であるから、水流停止の反応時間は、ほとんどが刺激の伝導に要する時間であったことになる。つまり、刺激から水流が停止するまでの時間は、遅い刺激伝導に要する時間と、水流ポンプ機構が停止する短い反応時間(潜時)の和で決まっていることになる。停止反応の絶対不応期は約30秒で、それ以下の間隔で連続二刺激を与えると、単一刺激に対する反応と同様の反応しか生じなかった[挿図4]。
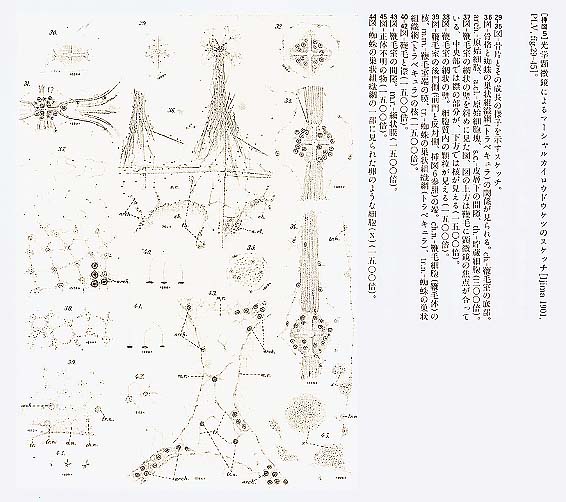
摂食水流を停止させる可能性のある機構としては、出水口、入水口、およびその間の水路が収縮して水流を妨害することも考えられる。ガラス海綿以外の海綿では、収縮による水路の閉鎖が観察されることがあるが、それは極めて遅い現象で、反応は全か無かの原則に従わず、ピークに達するまで30分程度も要するので、水流停止の機構としては時間がかかり過ぎる。しかも、ガラス海綿の堅い骨格から考えて、大きな部分が収縮によって変形する可能性は考えられないし、実際に水流が停止している間も、水路は広く開いたままであることが観察されている。また、ガラス海綿の表面を覆う皮膜(dermal membrane)を取り去っても水流停止反応が起るから、皮膜にある水流が通過する孔の収縮が水流停止の原因ではない。酢酸等の刺激物質や固定液を作用させても、皮膜の孔の直径は変化しない。ガラス海綿類に収縮能がないとすれば、この水流停止の機構は、水路の閉鎖ではなく、海綿全体に分布する襟細胞(襟鞭毛体)[挿図6参照]の鞭毛運動が一斉に停止することと推定される。ガラス海綿類以外の海綿では、水流の調節は、もっぱら収縮による水路の閉鎖によるとされている。従って、水流調節の機構は、ガラス海綿とそれ以外の海綿とでは、全く異なっていることになる。 海綿動物では、現在まで、鞭毛運動の停止が観察されたことは無いが、鞭毛や繊毛が刺激に応じて急激に逆転したり停止したりする現象は、単細胞生物のクラミドモナスの鞭毛やゾウリムシの繊毛等の繊毛逆転(ciliary reversal)や二枚貝やホヤ等の繊毛停止(ciliary arrest)等、多くの例で観察されている。バクテリアの鞭毛を除けば、鞭毛は繊毛と全く同じ細胞内小器官(organella)で、運動もその制御も共通の機構によっていると考えられている。そして、繊毛(鞭毛)運動では、その急激な反応は、膜の電気的な興奮(活動電位の発生)によって制御されていることが知られている。つまり、繊毛細胞では、興奮状態で運動を逆転ないし停止させ、平常状態で通常の方向に水を送る運動を継続するという制御が行なわれているのである。ガラス海綿では、その繊細な組織を傷つけると直ちに反応しなくなってしまうため、現在のところ、実際に鞭毛が停止するという観察は成功していない。実際に鞭毛が停止するのか、打ち方が乱れて有効に水を送ることができなくなるのかは判断できないが、ガラス海綿類においても、刺激に応じて鞭毛運動が停止または逆転している可能性は高いと考えられる。 5 ガラス海綿の刺激伝導系それでは、このガラス海綿の中で刺激を伝播させている機構はどのようなものであろうか。ガラス海綿の体のどの部分に触れても、水流の停止が起こる。外壁からでた一本の棘を動かすだけで水流は停止する。自然状態では、多数の顆粒が流入したときに、水流の停止が起こる。停止反応は、水温が摂氏15度以上になれば起こらなくなる。光には感じない。個々の襟鞭毛体が直接刺激に反応するかどうかは、わかっていない。電気刺激に対する反応の閾値は、標本によって非常に異なっている。しかし、敏感な標本では閾値は一定している。例えば2.6ボルトでは何の反応も起こらず、2.7ボルトでは完全な反応が起こり、水流の停止は全か無かの原則に従っていた。反応は、静かな流水に数時間そっとしておいたものに容易に起こり、水槽を灌流している水流を止めたり、強い機械的な刺激を与えたりすれば、起こり難くなった。このような標本に強い電気刺激を与えると、正常な停止反応が生じた。この結果は、閾値が高くなったのは、実行器である鞭毛の反応性の低下ではなく、刺激伝導系の興奮性が低下したためであることが示唆される。また、繰り返し機械的刺激を与え、反応性が低下した標本を数時間静置すれば、正常な反応が回復する。自然状態でも潮流等により繰り返し刺激が与えられる機会が多いことを考慮すれば、反応性の低下は、一種の適応現象(慣れ)であることも考えられる。 摘出した体壁で測定した伝導速度は、平均毎秒0.26(0.26±0.07)センチメートル(摂氏11度)で、時間的にも変動せず、場所や伝導方向にもよらず、ほぼ一定の価が得られた。被膜のみを残した部分の伝導速度は毎秒0.13(0.13±0.02)センチメートルと遅く、変動も大きかった。また、ジグザグに切った体壁の一端を刺激しても、興奮はその部分を越えて伝導した。興奮の伝導は、幅1.5ミリメートル、長さ20ミリメートルの大きさに切り出した体壁でも起こった。皮層と胃層に分けた標本では、その両者で伝導が見られたので、伝導系がどちらかの層に偏在することはない。静置した標本では、自発的な水流の停止が生じたが、周期性や概日リズムは認められなかった。繰り返して刺激された標本や、飼育条件がやや低下した状況では、次回に停止反応を生じたときには、一定の周期で繰り返し停止反応が起こる傾向が認められた。これは、ホヤや二枚貝の鯉でもよく観察される現象である。また、出芽によって二つの出水口を持ったものでも、繰り返し反応が見られたが、それぞれの口で記録された水流の停止は、時によって、その立ち上がりが前後し、それぞれ別のぺースメーカーがあるが、片方で生じた興奮は、自由に他方まで伝わってゆくことが示唆された。以上の結果は、刺激伝導系が海綿全体に均等に分布していることを示唆するものである。伝導の方向性も認められなかった。 伝導の機構としては、化学物質が刺激された箇所から放出され、その拡散によって反応が伝播する可能性も考えられる。しかし、反応性の良い二個体をとり、一方の出水口を他方の体壁に密着させて前者を刺激し、水流を停止させても、後者には反応は起こらなかった。また、体壁を切り出して二分し、それを密着させて片側を刺激しても、他方には反応が生じなかった。これは、物質の拡散や、水流停止の力学的影響で刺激が伝播する可能性が低いことを暗示する結果である。力学的な干渉を介して水流停止反応が伝播する可能性が低いことは、例えば、皮層のような、鞭毛の生えていない場所を通しても、刺激が伝導したという結果からも示されている。ガラス海綿以外の海綿類では収縮の波が伝播することが観察されている。しかし、この波の伝導速度は、毎秒8.30マイクロメータで、ガラス海綿の、毎秒2ミリメートルを越える停止反応の伝導速度にはとてもおよばないものである。細胞内を移動する化学物質が刺激を伝導する可能性は、最も速い軸索輸送でも毎秒2マイクロメータ以下で、ガラス海綿の刺激伝導速度の千分の一にも達しないことから、可能性は無いものと考えられる。 一方、活動電位は、神経細胞、筋細胞のような興奮性細胞だけではなく、ヒドロゾア等の上皮組織でも発生し、伝導することが知られている。ガラス海綿における伝導が全か無かの原則に従うことを考慮すれば、電気的伝導が、最も可能性が高いと考えられるが、傷つきやすい細い糸状の組織や、無数の孔があいた厚さ1マイクロメータの皮膜から、海水中で電気的活動を記録することは難しく、1983年当時、技術的にこれを証明することはできなかった。興奮の伝導機構を検討するためにも、ガラス海綿類の組織を、改めて詳しく調べる必要があったのである。 6 ガラス海綿の電子顕微鏡による観察飯島は光学顕微鏡による研究によって、ガラス海綿には、連続したシンシチウムの組織が網状に拡がっていることを報告した[挿図5、Ijima 1901]。その後、発生や再生時の組織を光学顕微鏡で観察すると、ガラス海綿類の組織が細胞膜の仕切りのない多核体より形成されてくること等、飯島の観察を裏付ける報告がなされていたが、マッキーらは、電子顕微鏡を用いてそれを確認すると共に、多くの興味深い観察結果を報告したのである。彼らは、潜水により得た完全で新鮮な標本を特に工夫した方法で固定し、ガラス海綿類の組織には原則として細胞間の仕切りがなく、個体が連続した細胞質でできた真の多核体であることを確認した[Mackie and Singla 1983]。
ガラス海綿類の組織は、基本的には、網のような細胞質のシート(網状膜 reticular membrane)からできており、全体は蜘蛛の巣状のトラベキュラ・シンシチウム(trabecular syncytium)または柱梁組織網(trabecular network)で繋がっている。鞭毛室の壁は全体として、連絡した2枚の網状膜が重なったもので、その上流側の膜の内面に鞭毛がある[挿図6]。ガラス海綿類には、海綿類の特徴とされる襟細胞は無く、それに似た襟と鞭毛を組み合わせた構造がある。その襟鞭毛体(collar bodies)は、多くの穴(前門賃 prosophyle)のある連続したシンシチウム(襟シンシチウム choanosynsytium)の、網の結び目に当たる場所についている。各襟鞭毛体に対応した核はない。襟鞭毛体の並んだ網状膜は、襟シンシチウムとトラベキュラ・シンシチウムが絡み合い、コラーゲン様物質でできたシート(メソラメラ mesolamella)に裏打ちされたものである。両シンシチウム間には細胞質の連絡が見られる。それらのシンシチウムの成因は、発生時や組織を再構成させた時に、まず多核の巨大細胞が形成されるという観察から、各細胞の融合したものではなく、むしろ、細胞質分裂を伴わない核だけの分裂により生じたものであることが示唆されている。各襟鞭毛体の間は連続した細胞質(ストロン stron)で連絡され、紅藻類に特有の細胞間連絡(壁孔連絡 pitplug)に似た、孔のある特殊な隔壁で仕切られている。その隔壁は、ゴルジ装置から同時にいくつも作られ、円盤状になってから、網目の細かい部分に送られてはめ込まれる[挿図7]。円盤は全体の厚みは約55ナノメータで、三層の電子密度の高い部分からなっているが、単位膜に相当する構造は見られず、ギャップ結合を始め、細胞膜が接して形成される接着構造とは似ていない。このような細部の構造は、もちろん光学顕微鏡では見ることができなかったが、連続した網目状の構造がシンシチウムであるという本質的な判断をした飯島の観察力には、敬服する外はない。飯島は、細胞質連絡に、何らかのバリヤーがあることも、その前後で染色性穎粒の濃度差があることから推測している。
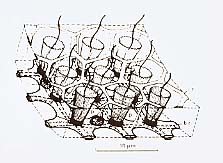
上記の隔壁は、生骨細胞(骨片母細胞 scleroblasts)以外の全ての細胞に見られている。ガラス海綿類の骨格は、多核の生骨細胞により細胞内で作られる。生骨細胞だけは、異種の細胞間で、直接連絡したり、隔壁で繋がったりすることはない。ガラス海綿類には、網状膜、トラベキュラ・シンシチウムの他に直径17マイクロメータにも達する貯蔵細胞(thesocyte)、直径3—5マイクロメータの穎粒が多い原始細胞(archaeocyte)等の細胞が観察されている。ガラス海綿類の細胞間には、通常の意味での細胞間の仕切りがないため、改めて細胞の定義を考えなければならないということはあるが、通常の膜で囲まれた細胞で構成されているガラス海綿以外の海綿類に見られる細胞の種類に、それぞれ対応した部分が見られることは興味深い。ガラス海綿類を含む海綿類には、細胞間の電気的結合に役立つといわれているギャップ結合(gap junction, nexus)は、見つかっていない。中隔接合(septate junction)は、トラベキュラ・シンシチウムや貯蔵細胞、原始細胞や襟鞭毛体等の異なる細胞種の間に、普通に見られる。原始細胞は、恐らく幹細胞で、そこからほかの細胞が分化して来ることが示唆されている。顆粒細胞は、直径5から8マイクロメータの電子密度の高い顆粒を多く含む細胞で、その機能は不明である。 飯島が、蜘蛛の巣状網(cobwebs)と表現したトラベキュラ・シンシチウムは、ガラス海綿の表面を覆う皮膜に連続し、典型的には厚さ1—2.5マイクロメータの薄膜になっている。膜には、直径4—30マイクロメータの多数の孔が開いている。一般の海綿動物に見られる扁平細胞(pinacocyte)は、ガラス海綿にはない。一般の海綿類の扁平細胞による皮層(dermal layer)の入水孔は、収縮して小さくなったり、場合により完全に閉じてしまうことが知られている。しかし、ガラス海綿の被膜の孔は、刺激しても大きさが変化せず、収縮能を示唆するような微細構造もない。それは、むしろ一般のトラベキュラ・シンシチウムと共通した内部構造を持っていることが確認されている[Mackie and singla 1983]。他の組織を含め、ガラス海綿類には収縮に関係すると考えられる繊維構造は観察されない。このことからも、ガラス海綿類の水流停止反応に、収縮機構が関係している可能性はほとんどないと考えられる。 7 活動電位の記録1997年になって、マッキーらは、上記のガラス海綿から、活動電位を記録することに成功した[Leys and Mackie 1997]。これは、広く海綿動物から電気的活動が記録された最初の例である。彼らは、ガラス海綿の組織を切りだして培養し、組織塊を作った。組織には、コンカナバリンAを作用させて凝集しやすくした。できた組織塊を、もとの海綿から切りだした板状の体壁の胃膜に接着させた。24時間後には、移植片は体壁と連絡し、細胞質が、体壁の組織に流入していくのが観察された。そのような標本の移植片に直径80マイクロメータの細胞外電極を吸い付け、その近くに水流計を設置した。記録電極から1センチメートルほど離れた場所を電気刺激すると、誘発された電気的活動とそれに続く水流の停止が記録された[挿図8]。電位変動は、二相性で、振幅は50—200マイクロボルト、持続時間は約5秒であった。この活動電位の伝導速度は、摂氏11度で、毎秒1.7—3ミリメートルであった。この電気的活動には30—40秒の不応期があったが、第一の刺激から40秒後に与えた刺激には、第一刺激と同様の反応が見られた。
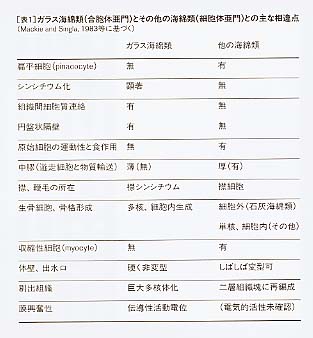
この結果は、ガラス海綿類には、電気刺激に応じて、伝導性の活動電位を発生する興奮性膜があることを示している。この性質をもった組織は、海綿の有機物質の75パーセントを占めているトラベキュラ・シンシチウムであると考えられる。この組織は、鞭毛室を含めて、ガラス海綿のあらゆる場所に連続した網となって連絡している。この網を通って伝えられた刺激が、さらに連続した襟シンシチウムの膜を興奮させることによって鞭毛運動を停止させるとする説には、確かな根拠が得られたと考えられる。マッキーらは、神経系の「網状説」が想定したような生物が現に存在することを示したのである。この膜の電気的活動が、どのようなイオン種のチャネルに関係しているかはわかっていない。この実験から、神経細胞のないガラス海綿類にも興奮伝導の機構があることが示された。これは、多細胞生物の進化の上で、膜興奮は神経細胞の出現に先立って生じたことを示唆している。 8 ガラス海綿類の系統について海綿動物の分類は、歴史的には石灰海綿類(Calcarea)とそれ以外の無石灰海綿類(Non-calcarea)に大きく二分し、それを六方海綿類と普通海綿類(Demospongiea)の二綱に分けて考えてきた。これは石灰海綿類が炭酸カルシウムを主成分とする骨片を持つという、はっきりした特徴を持ち、他のものと区別しやすいということを重視したものである。しかし、ガラス海綿類は、他の海綿類からばかりでなく、多細胞動物全体からも異なる独特の性質を持つことがわかってきた。マッキーらは、その組織を構築する基本的なプラン、特に細胞間の連絡の様式は、むしろ植物のものに近いことを示唆している。そして、ガラス海綿類の進化が、動物というよりはむしろ植物に近い方向をたどってきたように見えることを指摘している。海綿動物そのものが、幾度か、植物との類似性が指摘され、ある場合には、植物として分類されてきたことにも、それなりの背景があったと言えるかもしれない。 彼らは、襟鞭毛細胞と共通の襟鞭毛体を持つことに重点を置き、ガラス海綿類とその他の海綿類をまとめて海綿動物門とし、ガラス海綿類を合胞体亜門(Subphylum Symplasma)、その他の海綿類を細胞体亜門(Subphylum Cellularia)と二分して扱うことを提案している[Reiswig and Mackie 1983]。ガラス海綿類が、その他の海綿類と異なる主な点を表1にまとめた。
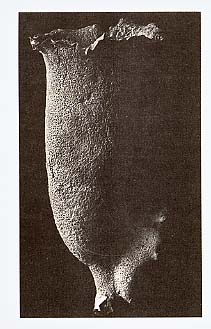
飯島の指摘したガラス海綿類の組織の特徴である細胞質の自由な連絡は、マッキーらの努力により確認され、ガラス海綿のほぼ全身が、細胞質の連絡したシンシチウムであることが示された。 広く動物界全般に分布している細胞間連絡の様式は、ギャップ結合である。ギャップ結合によって、小さな分子の細胞間の選択的な受け渡しが可能となり、物質の透過性が局所的な環境の変化によって調節できることとなった。それは、組織ごとに独自の機能を発揮させる可能性を大きく拡げさせ、また、膜興奮性を獲得した細胞組織では、イオンを通過させることによって、活動電位の伝播が可能となった。細胞問のギャップ結合の獲得は、多細胞生物の進化の上で、決定的に大きな役割を果たしたと考えられる[Mackie and Singla 1983]。しかし、植物とガラス海綿では、ギャップ結合が進化しなかった。そのため、その両者には、細胞質の連絡や、より不完全な細胞間隔壁が用いられている。ガラス海綿以外の海綿類に至っては、ギャップ結合はもとより、それに代わる何らかの細胞間結合も、獲得できなかったものと思われる。 水流停止がガラス海綿にとってどのような意義を持っているのかは現状では分からない。マッキーらは、他の一般的な例から、停止反応は一種の逃避反応であることを示唆している。筆者は、二枚貝の鰓や多くの海産無脊椎動物の幼生などに見られる例から考えて、多数の顆粒が入ったとき、それを効率良く捕食するために、一時的に水流を停止させる摂食行動の可能性が高いと考えている[Murakami & Machemer 1982]。 9 まとめ古典的な常識に従えば、海綿動物には、神経や筋肉は分化しておらず、最も原始的な動物群であるとされている。しかし、ガラス海綿類が、伝導系の起源ともいうべき機構を全身に張り巡らしているという事実は、私達の先入観を、改めて問い直すきっかけを与えてくれる。神経の興奮伝導は、生物の活性の中でも目覚ましい現象であるために、生命活動の本質的な機能の一つと考えられがちである。しかし、多細胞生物の進化の経過を想像すると、細胞問情報伝達の機構は、一朝一夕に完成されたものではなかったことが読み取れる。興奮性細胞の早い情報伝導の機構は、生物の歴史の上では比較的遅く進化した、むしろ特殊化した機能であると考えることもできる。人間中心の考え方では、神経活動こそが最も生物らしい機能の現われであり、全ての動物の進化の到達点が、人の大脳のような機能を獲得することであると考えがちである。無脊椎動物学の知見は、このような発想を修正し、生物の多様な在り方を客観的に把握する基盤を与えてくれるものと考える。また、ガラス海綿の研究成果は、私達に、動物と植物の関係に新しい見方を提供してくれた。膜の電気的活性によって運動を制御している例は、植物にも報告されている。生物の分類は、生物を動物と植物に分けて考えるところから始まった。そのため、私達は、動物と植物の違いは、実際以上に本質的なものであると錯覚しているのかもしれない。 ガラス海綿は、ほぼ全身が細胞質の連絡したシンシチウムである。ガラス海綿には、大きなものがあり、飯島の標本の中にも高さ88センチメートルに達するものがある(ツリガネカイメン[挿図9]。これは、現在知られている動物としては、最大のシンシチウムである。この動物全体が一つの原形質の塊で、縦横に刺激を伝え、全身が一度に興奮して、びっしりと並んだ鞭毛室の内面を覆う鞭毛が一斉に止る光景は、壮観を通り越して、怪奇ドラマの1シーンを思わせるものがある。
標本をじっと見つめ直したその日の印象を記して結びとしたい。標本は変わらない。棚の片隅に、ひっそりとたたずんでいる。しかし、ある日、突然それは輝くのである。その輝きをもたらしたものは、現実と幻影の逆転である。巨大な海綿の塊は、昨日までは、ばらばらな細胞の寄せ集まりの見本としてそこに居た。いかにもその表情は、遅れ馳せながらやがてギャップ結合を進化させ、まともな多細胞動物の仲間入りができる日を待っている、うつ向きかげんの控えめなものであった。しかし、今日の表情は違う。それは、消え去ったかに見えた網状説の生きた見本として輝いている。全身が細胞質の連絡した糸として繋がり、興奮を縦横に伝える生き物が、現実に存在していることを語りかけているのである。たとえ今はおぼろげな伝説としてしか知られていない古代の文明が、厳然として存在したことを示すインカ帝国の黄金の仮面のように、目の前に輝いているのである。もちろん、標本は変わらない。それを変えたのは、その見方を変えた人である。そして、標本は、自分の見方を変えてくれる人を待っている。今日も。 |


【参考文献】堀越増興「六放海綿類の研究—飯島魁とその業績—」、『研究総合資料館展示解説』、東京大学総合研究資料館、1983年、109—114頁。Ijima, I. Studies on the Hexactinellida. Contribution I. (Euplectellidae). J. Coll. Sci. Imp. Univ. Tokyo, 1901, 15, 1—299. Ijima, I. Studies on the Hexactinellida. Contribution IV. (Rossellidae). J. Coll. Sci. Imp. Univ. Tokyo, 1904, 18, 1—307. Lawn, I.D., Mackie. G.O. & Silver, G. Conduction system in a sponge. Science, N. Y., 1981 , 211, 1169—ll71. Leys S P & Mackie G.O. Electrical recording from a glass sponge. Nature 387(6628), 1997, 29—30. Mackie, G.O. & Singla. C. L. Studies on hexactinellid sponges. I. Histology of Rhahdocalyptus dawsoni (Lambe, 1873). Phil. Trans. R. Soc.Lond., 1983, B 301, 365—400. Mackie, G.O., Lawn I. D. & Pavans de Ceccatty, M. Studies on hexactinellid sponges. II. Excitability, conduction and coordination of responses in Rhabdocalyptus dawsoni (Lambe, 1873). Phil. Trans. R. Soc. Lond., 1983, B 301, 401—418. 萬年甫「光学顕微鏡でみた世界 総論」、黒川正則・塚田裕三・萬年甫編『神経生物学』、共立出版、1967年、20—37頁。 Murakami, A. & Machemer. H. Mechanoreception and signal transmission in the lateral ciliated cells on the gill of Mytilus. J. Comp. Physiol., 1982, 145: 351—362. 村上彰・富永義昭「六放海綿類と日本産魚類 展示紹介—動物部門」、『総合研究資料館ニュース』、東京大学総合研究資料館、1986年、4—5頁。 Reiswig, H. M. & Mackie. G.O. Studies on hexactinellid sponges. III. The taxonomic status of Hexactinellida within the Porifera. Phil. Trans. R. Soc. Lond., 1983, B 301, 419—428. |
[東京帝国大学理学部動物学科]
150 昆虫学レファレンス・コレクション(箕作佳吉収集) 乾燥標本約5千点、明治17(1884)年—明治38(1905)年、総合研究博物館動物部門 明治14(1881)年東京大学理学部動物学教室の教授となった箕作佳吉の収集になる昆虫標本。コウチュウ目20箱、チョウ目33箱、その他3箱からなり、全体として5千点を超える昆虫類が保存されている。ドイツ人フリッツェが北海道で採集したオオキノコムシをはじめ、ギフチョウの発見者として知られる名和靖が明治22年に採集した「ギフヤマ」、中川久知の採集したシジミチョウ類、高千穂宣麿が明治17年に採集したエルタテハなど、歴史的に意義深い標本が含まれている。また標本箱のなかには箕作がチョウ類の擬態に関する講義に用いたとみられるペン書きのメモ類も残されている。現在の学制では昆虫学が農学系諸学の一分野と見なされているが、箕作の時代には理学部動物学教室に属していたことを物語る史料である。なお、箕作は明治26年のシカゴ世界博覧会にも日本産昆虫標本を出品しており、ワシントンの国立博物館にそれらがそのまま保存されている。(西野)
151 動物学レファレンス・コレクション 液浸・乾燥標本、明治後期、総合研究博物館動物部門 151-30 オキクラゲ Pelagia noctiluca 液浸標本、高18.5cm、径9.0cm、「Naples」の記載あり 151-46 コウイカ類の卵目 Egg of Sopia 液浸標本、高10.0cm、径3.5cm 151-50 キヌタアゲマキ Solecurtus divaricatus 液浸標本、高20.0cm、径6.5cm 151-54 シャミセンガイ類の一種 Lingula anatina(二点) 液浸標本、高10.0cm、径3.5cm/高13.5cm、径5.0cm、「Okayama」の記載あり 151-71 コオオグソクムシ Bathynomus doederleini、ウオジラミ Bathynomus doederleini Ort.(?) 液浸標本、高22.0cm、径9.5cm、「下浦」の記載あり 151-72 カメノテ Mitella mitella 液浸標本、高10.5cm、径3.5cm、「対馬□原」の記載あり 151-79 ガンガゼ Diadema setosum 液浸標本、高12.0cm、径6.0cm、「小□房州」の記載あり 151-83 ヒメウミシダ類の一種』Antedon inequalis 液浸標本、高19.0cm、径6.0cm、「Misaki」の記載あり 151-114 ヨーロッパ産のアカガエルの一種 Rana esculenta 液浸標本、縦43.0cm、横9.0cm 151-98 ヘラチョウザメ Polyodon spathulan(二点) 乾燥標本、長141.8cm、高38.2cm、幅33.2cm/長128.4cm、高23.1cm、幅21.5cm 151-101 フグ類の一種 乾燥標本、長71.2cm、高24.8cm、幅21.2cm 151-113 カイメン(箱入) 乾燥標本、長58.5cm、高39.5cm、幅37.0cm 151-115 パイプウニ、Heterocentrotus momillatus 液浸標本、縦15.5cm、横12.0cm 151-116 ツッイカ類の一種 液浸標本、縦12.5cm、横4.5cm 151-117 ヤツメウナギ類の一種 液浸標本、縦20.5cm、横9.0cm |



| 前頁へ | 表紙に戻る | 次頁へ |