Phytogeography of the Qinling Mountains and a Comparison with the Flora and Vegetation of Japan
Tsun-shen Ying1 and David E. Boufford2
1Institute of Botany, Chinese Academy of Sciences, Nanxincun 20, Xiangshan, Beijing 100093
People's Republic of China
2Harvard University Herbaria, 22 Divinity Avenue, Cambridge, Massachusetts 02138, U.S.A.
|
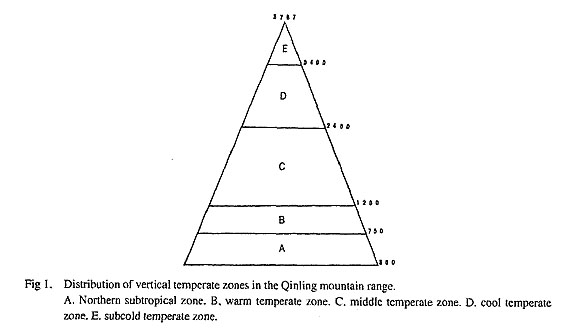
Abstract. The Qinling Mountains, a major watershed of the Huang He (Yellow) and Chang Jiang (Yangtze) rivers, cover an area ofca. 76,500 km2 and range from 32°05' to 34°45'N and from 104°30' to 115° 52'E. The flora of the area comprises ca. 3,125 species of flowering plants in 892 genera, of which 39 of the latter are endemic to China. The significance of the 20 largest families, which together contain 65.2% of the total species, is evaluated, and the phytogeographical affinities of the genera of spermatophytes in the flora are analyzed and briefly discussed. In the flora of the Qinling Mountains 63.8% of the genera also occur in Japan. The distributional patterns of species are analyzed and some comparisons are made with the flora of Japan. Endemic species and non-endemic species are nearly equally represented in the Qinling mountain range; 49.1% of the non-endemic species are shared with Japan. Analysis of the composition of the different forests makes it clear that the vegetation can be thought of as comprising two main elements: an endemic Chinese element and a Sino-Japanese element. Key words. Phytogeography, Japan, China, Qinling Mountains, endemism, geographical limits, soil types and climate. The Qinling Mountains, which occupy an area of ca. 76,500 km2 and reach from 32°05' to 34°45'N and from 104°30' to 115° 52'E, constitute a major watershed of the Huang He (Yellow) and Chang Jiang (Yangtze) rivers and form an important and conspicuous geographical divide between northern and southern China. The highest peak, Taibai Shan, is 3,767 m above sea level. The relative elevation in the western part of the range varies from 2,000 to 3,000 m; most of the eastern part is below 2,000 m. Five major soil types have been described from the Qinling Mountain region: 1) ochric castanozom below 1,000 m; 2) brown forest soil between 1,000 and 1,500 m; 3) podzolic brown earth from 1,500 to 2,800 m; 4) podzolic soil from 2,800 to 3,000 m; and 5) alpine meadow soil above 3,000 m (Ying et al., 1990). An analysis of the climatic features of the Qinling mountain range was made by Chen (1992), who classified the vertical climatic zones from lowest to highest elevation as northern subtropical, warm temperate, middle temperate, cooler temperate and subcold temperate (Fig. 1). When the temperature and humidity of the south and north slopes of the Qinling Mountains are compared, significant differences are observed. On the south slope, the average annual temperature is 2°C higher than on the north slope, the winter average temperature is 4°C to 5°C higher, the relative humidity is 10% higher and the average rainfall is 200 to 400 mm greater.
|
|

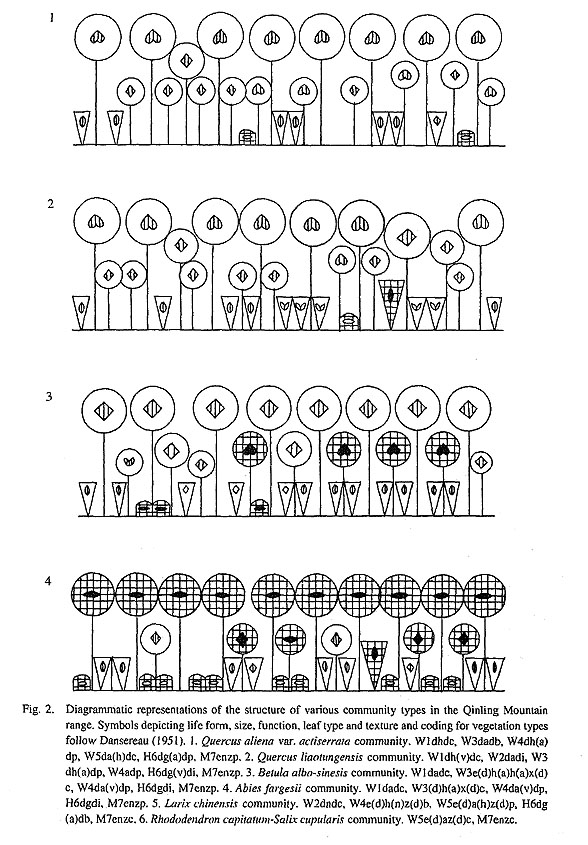
There are also many endemic species in the composition of the plant communities of the Qinling mountain range. For example, 126 of the 217 species within the Quercus aliena var. acutiserrata forests are endemic to China, including seven restricted to the Qinling mountain range. One hundred sixteen of the 184 species in the Quercus liaotwigensis communities are endemic to China and six occur only in the Qinling mountain range. The Betula albo-sinensis forests contain 152 species, of which 111 species are endemic to China, and five are endemic to the Qinling mountain range. The Abies fargesii forests contain 105 species, 72 of which are endemic to China, including seven restricted to the Qinling mountain range. The Larix chinensis forests contain 114 species, 76 of which are endemic to China, including 11 endemic to the Qinling mountain range.
Antiquity appears to be another outstanding characteristic of the forests in the Qinling Mountains. Carpinus cordata Blume, of common occurrence in the Qinling Mountains, is morphologically close to the fossil species C. subcordata, while another common species C. fargesii Franchet is close to the fossil species C. miofargesiana (Tanai, 1972). Fossil species of Carpinus L. are also found in Tertiary deposits of North America and Europe, Fossils attributed to the extant Lonicera hispida Pallas ex Roemer & Schulte have been found in a Miocene deposit in Shandong Province (Institute of Botany and Nanjing Institute of Geology and Paleontology, Academia Sinica, 1978). The dominant species Quercus variabilis is morpho-logically similar to the fossil species Q. miovariabilis, which was also deseribed from the Miocene of Shandong (Hu and Chaney, 1940). Another dominant species Quercus liaotungensis is remarkably similar to the fossil species Q. cf. liaotungensis which was found in Miocene deposits in Shanxi Province (Institute of Botany and Nanjing Institute of Geology and Paleontology, Academia Sinica, 1978). The occurrence of extinct and extant species within a genus in the same geographical area may also indicate the antiquity of the community in which the living species occur, although it is hard to imagine that these plants remained stationary during the climatic fluctuations of the Pleistocene.
Temperate forest characteristics are another prominent feature of the vegetation of the Qinling mountain range. As shown in Figure 2, except for Abies fargesii all other forest dominants are deciduous trees. There are no evergreen broadleaved forests, or even mixed evergreen-deciduous broadleaved forests. The forests are composed of about 526 species belonging to 261 genera and 92 families. Except for Elaeagnus L. (2 spp.), Ilex L. (1 sp.). Daphne L. sp.). Rhododendron L. (3 spp.) and several species of conifers, almost all of the other species are deciduous. In addition, most of these genera and species are East Asiatic elements and indicate a very close relationship between the flora of the Qinling mountain range and the flora of Japan, especially the flora of central Japan.
Floristic relationships manifested through the deciduous forests of the Qinling mountain range and Japan
Broadleaved deciduous forests prevail in the north temperate zone and are particularly well developed in China, Japan, eastern North America, Europe and the Caucasus Mountains (Daubenmire, 1978; Numata, 1974; Wu, 1980). Due to similarities in geographical characteristics and climatic conditions, and no doubt because of their close proximity, the composition of the vegetation in the deciduous forests of China and Japan is more similar than it is between China and eastern North America, Europe and the Caucasus. The dominant and/or common species of the broadleaved deciduous forests of China and Japan, especially those in central China and central Japan, are often members of species pairs or are closely related species of the same genus, as shown in Table 1 (see also Cheng, 1984). The relationships between the Qinling mountain range and Japan are not limited to the dominant and common species. The floristic composition of the deciduous forests also contain many species occurring in both regions. The Quercus aliena var. aculiser-rata forest contains 217 species, of which 126 species are endemic to China. The remaining species are mostly conspecific with the members of the Japanese flora. The Quercus liaotungensis forests contains 184 species in 118 genera and 53 families. Among them 116 species are endemic to China; 55 of the non-endemic species have an eastern Asian distribution. In the Betula albo-sinensis forests are 152 species, including 111 endemics, and 41 non-endemics of which 26 occur in Japan. The B. utilis forests contain 173 species, including 119 endemics and 54 non-endemics; 33 of the latter are also in Japan. These non-endemic species manifest some of the characteristics common to the flora of the Qinling mountain range and central Japan.
 
|
Survey of the largest families of angiosperms
The largest families of flowering plants in the Qinling mountain range are the Asteraceae (355 species), Poaceae (219 species), and Rosaceae (199 species) (Table 2). The Rosaceae are a major family in this region and are particularly characteristic of the flora and vegetation of temperate areas in China. The Asteraceae and Poaceae also exhibit an extraordinary frequency in the forests, and elsewhere in this region. There are seven families with more than 100 species: the Fabaceae (175 species), Ranunculaceae (128 species), Cyperaceae (121 species), Lamiaceae (116 species), Apiaceae (110 species), Liliaceae (103 species), and Orchidaceae (102 species). The development of the Fabaceae is mainly in the Papilionoideae, which are represented by 166 species; the Mimosoideae and Caesalpinoideae have only three and six species respectively. The Ranunculaceae and Cyperaceae are north temperate in nature and form the major part of the flora. The Lamiaceae and Apiaceae comprise an important element of the flora, and three genera of Lamiaceae endemic to China occur in the Qinling mountain range. Two families, the Liliaceae and Orchidaceae, are comparatively poorly represented, but two genera and 50 species of Orchidaceae endemic to China occur here. The ten families mentioned above comprise nearly 48.3% of the species of flowering plants in the Qinling mountain range. There are ten additional families with between 40 and 100 species. In order of size they are: Caprifoliaceae (74 species), Salicaceae (73 species), Scrophulariaceae (66 species), Saxifragaceae (65 species), Polygonaceae (58 species), Brassicaceae (57 species), Oleaceae (47 species), Berberidaceae (44 species), Caryophyllaceae (41 species), and Rubiaceae (41 species). Except for the Oleaceae and Rubiaceae, these families are north and south temperate, or only north temperate, in distribution. In addition, the Berberidaceae and Saxifragaceae contain 44 and 47 species respectively endemic to China, and have the highest proportion of endemic species of all of the larger families. The families mentioned above contain a combined total of 2,193 species, including 1,041 species endemic to China, or 65.2% of the total flora, and play an important role in shaping the characteristics of the flora and vegetation of the Qinling mountain range.

Relationships of the Qinling mountain range flora
A. Geographical affinities of the genera
Approximately 892 genera and 3,125 species occur in the Qinling mountain range. Based on the geographical distribution given for each genus in Willis's Dictionary of the Flowering Plants and Ferns (Shaw, 1973), Mabberley's (1987) The Plant-book, and Wu's (1991) Areal-types of Chinese Genera of seed Plants,we were able to classify the genera of flowering plants in the Qinling mountain range into 15 distribution types (Table 3) as follows.
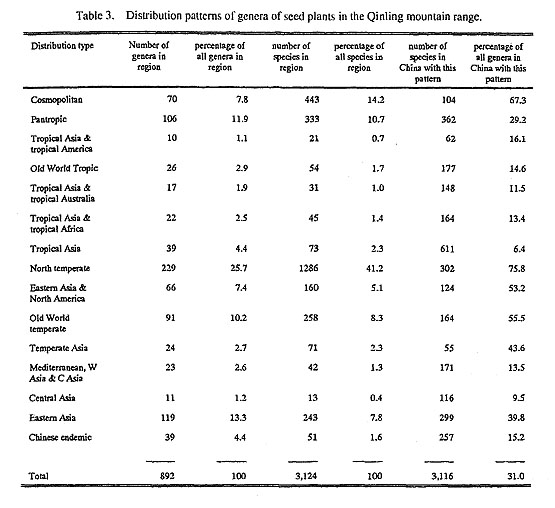
1. Cosmopolitan genera. Some genera, mainly aquatics and mesophytes, are so widespread that they can be classified as cosmopolitan. Seventy genera fall into this category. These taxa belong to 36 of the 158 native families in the Qinling mountain range and include 443 species. These cosmopolitans comprise about 14.2% of the flora of the Qinling mountain range, and 22.8% of the total families. Many mesophytes such as Carex L., Polygonum L., Ranunculus L., Viola L., Gentiana L., and Cyperus L. are either the dominant or common species of the herbaceous layer in forests and in montane grasslands.
2. Pantropical genera. Genera found in all three tropical regions (the Americas, Africa-Madagascar, Asia-Australasia) may be considered pantropical. In the Qinling mountain range 106 genera, or approxi-mately 11.9% of the total genera, may be classified as pantropical. They contain 333 species, or 10.7%, of all species in the flora of the Qinling mountain range. Although the centers of distribution of these genera are primarily concentrated in the tropics of both hemispheres, some of them, such as Ilex L., Cynanchum L., Impatiens L., Buddleja L., Ficus L., Cuscuta L. and Pilea Lindley, extend to subtropical or even temperate regions. It should be mentioned, however, that the distributions of many of these genera in the tropics are often in cool, moist, montane areas. One example of a tropical genus that has its northern boundary in the Qinling mountain range is Aeschynomene L. The genus contains about 150 species distributed throughout the tropics of both hemispheres. Only a single species. A. indica L., occurs in southern, southwestern, eastern and central China.
3. Tropical America and tropical Asia. Only ten genera in nine families. representing 1.1 % of all genera, are included in this category. These genera. all of which are woody. contain 20 species, or only 0.6% of the total flora of the Qinling mountain range. These genera are usually distributed at lower elevations in the Qinling mountain range. but a few of them occur in forests at higher elevations. In China some of these genera, such as Meliosma Blume, Sagereia Brongn. and Picrasma Blume, may extend north of the Qinling mountain range. but most of them, such as Phoebe Nees. Sloanea L., Eurya Thunb. and Sapindus L. have their distribution entirely to the south of the Qinling Mountains.
4. Old World tropics. This distribution type is difficult to distinguish from the subtropical and sometimes even temperate pattern. An example is found in the genus Melia L.. which has about 15 species in the Old World tropics. but only two species. one of which extends as far north as the northern slope of the Qinling mountain range. in China In the Qinling mountain range 26 genera, or approximately 2.9% of the total genera, may be classified as Old World tropical. These genera contain about 54 species, or 1.7% of all species in the flora of the Qinling mountain range. Among them. Elatostema Gaudichaud, Cayratia Juss. and Asparagus L. are constituents of the herb layer of the broadleaved deciduous forests, while the shrub Alan-gium Lam. is also rather common, but only along streams or at the edges of forests.
5. Tropical Asia and tropical Australia. The western extremity of this distribution type extends to Mada-gascar. but not to the African mainland. An example is found in Balanophora Forster & G. Porster, of which 80 species occur in Australia. Polynesia, Malaysia, South China. Japan and Madagascar, but none occur on the African continent. Seventeen genera (1.9% of all genera) and 31 species (1.0% of all species) can be placed in this category. Among the genera with this pattern of distribution, most are woody. In China these genera occur predominantly in southern China and extend northward to the southern slope of the Qinling Mountains. Only a few. such as Leptopus Decne., Cudrania Trec., and Gastrodia R. Br., reach the northern slope. The genus Nothopanax Miquel has a disjunct distribution in southwest China and New Zealand. Some genera, such as Wikstroemia End]., Leptopus and Nothopanax are constituents of the forests, but are not common.
6. Tropical Asia and tropical Africa. Genera in this category are distributed from tropical Africa to the Indo-Malaysian region. but some genera extend to Fiji or other islands of the South Pacific, but not to Australia. A good example of this pattern is seen in the monotypic genus Toddalia Jussieu which extends from eastern Africa and Madagascar to the southern part of the Asian continent and islands. In China, Toddalia extends as far north as the south slope of the Qinling mountain range. In the Qinling range, 22 genera (2.5% of all genera) belong to this distribution type. These genera contain 45 species (1.4% of the total species). Genera belonging to this pattern of distribution, except for Miscanthus Andersson and Microstegium Nees, are uncommon in this region.
7. Tropical Asia. The Indo-Malesian region has one of the richest floras in the world. The eastern portion of the Indian region and the northwestern portion of the Malesian region, through the Malay and Indo-Chinese peninsulas, are very closely linked floristically with the mountains of south-central China. Thus, a fairly large number of "typical" Indo-Malesian genera occur in south China. For example, this category forms the nucleus of the flora of Guangdong Province. In the Qinling Mountains, however, this distribution type is represented by only 39 genera, or 4.4% of all genera. Together they contain about 73 species, which comprise only 2.3% of all species. The genus Gynostemma Blume is a typical representative of this distri-bution type. Gynostemma contains 13 species, one of which occurs on Timor Island; one in Kalimantan; four in China, India, Sri Lanka, Nepal. Sikkim. Burma. Thailand. Vietnam. Malaysia. the Philippines, Indo-nesia, Japan and Korea: another seven species are concentrated in southern and southwestern China. The northern boundary for the distribution of the genus is the southern slope of the Qinling Mountains.
8. North temperate. Included under this heading are all the widely distributed genera restricted to the temperate regions of Eurasia and America. Their distributional areas sometimes extend southward to the more mountainous regions of the tropics. For example, Juglans L. contains 23 species, of which five are distributed in Asia, one in Europe and Asia. and the remainder in America; five species occur in the tropical mountainous regions of South America (Lu. 1982). The genera belonging to this category total 229, or 25.7% of the total genera enumerated, and represent the highest proportion of distribution types in the flora of the Qinling mountan range. The genera in this category contain ca. 1,286 species, or over 41 % of all species in this region. and form the nucleus of the flora of the Qinling mountain range. Among the especially prominent and significant genera in terms of their dominance in the vegetation of this region are Abies Mill., Larix Mill., Betula L., Quercus L., Salix L., Rhododendron, Spiraea L., Thalictrum L., Polygonatum Mill., Pyrola L. and Allium L. Most of these genera contain only deciduous trees and shrubs, and all of them are typical North Temperate genera.
9. Eastern Asia and North America. This pattern has been one of the best known and has attracted the greatest interest by biogeographers since Asa Gray (1846) first drew special attention to it, and it has been much studied and discussed (Boufford and Spongberg. 1983; Graham, 1972 a, 1972 b; Hara, 1952, 1956, 1972; Hu, 1935, 1936; Li, 1952; Wu, 1983; Ying, 1983). In the Qinling mountain range 66 genera, which represent 7.4% of the all genera, belong to this distribution type. They contain 160 species, or 5.1% of all species in the region. These 66 genera also make up about 60% of the disjunct genera with distributions restricted to eastern Asia and temperate North American. Among them Diphylleia Michaux, Buckleya Torrey and Clintonia Raf. are typieal representatives of this distribution type. The former contains three widely disjunct species, with one each in Japan (and extending to Sakhalin Island), China, and in eastern North America (Ying et al., 1984). Clintonia contains five species; one in eastern Asia, two in eastern North America, and another two in western North America. Buckleya contains four species, the very rare B. disti-chophylla (Nuttall) Torrey in eastern North America, B. lanceolata (Sieb. & Zucc.) Miq. in Japan, and B. graebneri Diels and B. henryi Diels in China. The two species of the genus in China are centered in the Qinling Mountains and B. graebneri occurs barley, if at all, outside of that region.
Although genera with this distribution pattern are concentrated in eastern Asia and North America, some of them extend into central Asia (Veronicastrum Moench), or rarely to Australia (Lespedeza Michaux) in the Old World, and occasionally even into tropical regions in the New World (Muhlenbergia Schreber). In addition, the genus Abelia R. Br. is also rather common in this region, and is distributed disjunctly in eastern Asia and Mexico.
10. Old World temperate. In the flora of the Qinling mountain range, 91 genera, or 10.2% of all genera, containing 258 species, or about 8.3% of all species, are Old World temperate in distribution. Most of these genera are herbaceous and are the main constituents of montane grasslands or of the herb layer in forests.
11. Temperate Asia. Included here are all the genera distributed mainly in temperate Asia, but whose dis-tribution sometimes extends southward to the more elevated regions of the subtropical zone. For example, Trigonotis Steven. contains about 57 species, of which 34 are distributed in China, and almost all of which are concentrated in the subtropical regions of the country. These species show their temperate nature, how-ever, in their ecologically restricted occurrence to areas with elevations between 1,000 and 4,900 m. We have segregated this group of genera mainly on the basis of the ecological preference of their subtropical representatives. In the Qinling mountain range this distribution type is represented by 24 genera, or 2.7% of all genera, which together contain about 71 species, or 2.3% of all species. Excepting Caragana Fabr., Campylotropis Bunge, Exochorda Lindley and Armeniaca Mill. (=Prunus L. subg. Prunophora (Necker) Focke), all of these genera are herbaceous.
12. Mediterranean, western Asia to central Asia. In the flora of the Qinling mountain range, only 23 genera, or 2.6% of all genera, belong to this distribution type. They contain 42 species, or about 1.3% of the to tal. These genera usually display a disjunct distribution pattern. For example. Peganum L. is disjunctly dis-tributed from the Mediterranean through Mongolia to northwestern China and from the southern United Slates to Mexico. Except for Gypsophila L. and Erodium L'Herit.. all of the genera of this distribution type are uncommon in this region.
13. Central Asia. This type is represented by proportionally few genera, especially when compared with the North Temperate distribution type. In the flora of the Qinling mountain range this distribution type is represented by only 11 genera, or 1.2% of all genera. Together they contain 13 species, which comprise only 0.4% of all species, and all of them are herbaceous. Except for Notholirion hyacinthinum (E. H. Wilson) Stapf, which is the main constituent in the herb layer in forests of Abies fargesii, none of them are sig-nificant constiwents in the forests of this region.
14. Eastern Asia. Included under this heading are all the genera restricted essentially to China and Japan. This category is a large one calculated to contain 119 genera or 13.3% of all genera in the Qinling region. These genera contain 243 species, or about 7.8% of the total, which can be divided roughly into three sub-groups based on their distribution centers in relation to the Qinling mountain range. One subgroup includes genera extending from the Himalayan region across China to Japan; the genus Helwingia Willd. is a typical representative of this subgroup. The other two subgroups have eccentric distribution centers, either toward the west or to the northeast. Members with a westward distribution show a Sino-Himalayan range, from China to the Himalaya. Circaeaster Maxim. is a typical representative of this distribution subgroup. The northeastward group shows a Sino-Japanese distribution pattern, from China to Japan. Idesia Maxim. is a typical example of this distribution subgroup. Among the 119 Eastern Asian genera and 243 species with an East Asian distribution, about 46 genera and 121 species belong to the Sino-Himalayan-Japan subgroup, 41 genera, containing 56 species, belong to the Sino-Japanese subgroup, and 45 genera with 66 species be-long to the Sino-Himalayan subgroup. These genera sometimes extend southward into Burma, India, Viet-nam, and/or the Malay Archipelago.
15. Endemic genera. For the Qinling mountain range as a whole, 39 of the 892 native genera (4.4%) are endemie to China, but all of them are shared with other provinces. These endemic genera are listed with their ranges, life form and number of species in Table 4. Twenty of these genera are monotypic and 16 are oligotypic; the three remaining genera have multiple species. Among the 39 are 17 arborescent genera, of which 15 are deciduous and two are evergreen. Eighteen genera are herbaceous, of which four are annual and fourteen are perennials. The four remaining genera are lianoid. Except for Notopterygium Boissieu, Si-nacalia H. Robinson & Brettell and Nannoglottis Maxim., which occur on the Qinghai-Xizang plateau, and Pteroxygonum Dammer & Diels, Myripnois Bunge and Speranskia Baillon, which also occur in Inner Mongolia and/or in northeastern China, all are restricted to southeastern, central and southwestern China. Kingdonia Balf. f. & W. W. Sm. is one of the main constituents of the herb layer in the Abies fargesii forests.
 
|
Based on the figures cited above we were able to conclude that among the native genera in the flora of the Qinling mountain range, 220 (26.8%) are tropical, 563 (68.5%) are temperate, and 39 (4.7%) are genera restricted to China. It is clear from the numbers and percentages that the temperate genera are the most sig-nificant elements in the flora and vegetation of the Qinling mountain range. Of the 892 genera in this region, 653 (73.2%) are shared with Japan.
B. Analysis of the species
1. Endemic species. Of the approximately 3,124 native species in the flora of the Qinling Mountains about 1,620 (51.9%) are endemic to China. Of the 192 species restricted solely to the Qinling Mountains, 60 are woody, 128 are herbaceous, and four are lianoid. The woody species, except for Jasminum humile L. forma kansuense (Kobuski) Miao, are all deciduous. Among the 128 herbaceous species only four species (1.6%) are annual. The proportion of annual endemic species is indeed low in this region. Among these en-demic species, 40 endemic species are restricted to the western Qinling Mountains, 12 to the eastern part, and 136 to the central part, of which 29 extend westward to the western part and ten extend to the eastern part. Only four endemic species have a widespread distribution in the Qinling Mountain range. Most of these endemic species occur between 800 and 2.000 m elevation. In addition to the species that are strictly endemic to the Qinling mountain range, about 1.428 species in the flora of the Qinling mountains are en-demic to China. Among these endemic species 1,181 are dicots, 219 are monocots and 28 are gymnosperms.
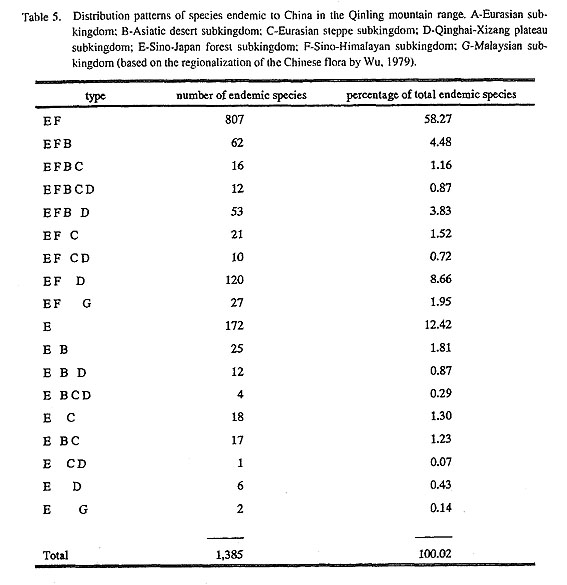
The division of the Chinese flora into distinctive regions was proposed by C. Y. Wu in 1979. He divided the Chinese flora into two kingdoms, seven subkingdoms and 22 regions. Based on their distribution among the seven subkingdoms, the endemic species discussed above can be classified into two types, eight subtypes and eight groups. As shown in Table 4, 807 endemic species are distributed in subkingdoms E and F. Among these species 11 are gymnosperms, 63 are monocots and the remaining species are dicots. The families and genera to which these species belong are all temperate in nature. About 62 species extend northwestward to subkingdom B and form an E-F-B distribution pattern. Twenty one species extend north-ward through Inner Mongolia into northeastern China to subkingdom C and form the pattern E-F-C. One hundred twenty species extend to the west, forming the E-F-D pattern. Only 29 species are distributed southward to subkingdom G to form the E-F-G distribution pattern. These species belong to 27 genera, all of which, with the exception of Glochidion Forster & G. Forster, Buxus L., Ilex, Sycopsis Oliver, Pleione D. Don and Amentotaxus Pilger, are temperate in distribution. As show in Table 2, it is also clear that ex-cept for Orchidaceae, Fabaceae and Oleaceae, all families are primarily temperate, north temperate or cos-mopolitan in their distribution. Approximately 172 species are entirely restricted to subkingdom E. These species are all temperate, and are the major constituents of the flora and vegetation in the Qinling Mountains. About 25 species extend northward to B, and 18 extended northeastward to C. They form E-B and E-C distribution patterns. Only six species extend westward to D and two extended southward to G. They be-long to the E-D and E-G patterns. Those endemic species restricted to subkingdoms E-F and E not only in-dicate that the flora of the Qinling mountain range comprises the main body of the Sino-Japanese forest subkingdom, they also provide reliable evidence of the floristic relationships between China and Japan.
2. Non-endemic species. Of the approximately 3,127 native species in the Qinling mountain range about 1,505 (ca. 48.1%) are non-endemic species. Among these species about 39 species are shared with Korea and about 739, or 49.1% of the total non-endemic species, occur in the flora of Japan. In addition, Cephalo-taxaceae, Eupteleaceae and Cercidiphyllaceae are of Sino-Japanese distribution. The Cercidiphyllaceae are represented by two species in the single genus Cercidiphyllum Sieb. & Zucc. Both species are in Japan and one of those species also occurs disjunctly in central China. As already mentioned in discussing the geo-graphical affinities of the genera, the flora of the Qinling Mountains has a high number (13.3%) of eastem Asian genera. It is clear in the numbers and percentages that the flora of the Qinling mountain range has closer phytogeographical links with the flora of Japan than with any of the other surrounding regions, in-cluding western China. Approximately 98 species occur in Siberia and the Russian Far East. Fifteen species are distributed northward into Mongolia and 54 are in the former Soviet Union. All of these species are temperate in nature. Of the approximately 1.505 non-endemic species about 340 extend to subtropical or even tropical areas. They occur in India (151 species), Burma (59 species), Indochina (41 species), the Phil-ippines (41 species), Malaysia (131 species), and Indonesia (17 species). For most of these species the south slope of the Qiniing mountain range is their northern boundary. For example, Lonicera acuminata Wallich is concentrated in south China and extends southward to Burma, the Philippines, Sumatra and Java. In China its northern boundary is on the southern slope of the Qinling mountain range.
Composition of the Vegetation
1. Composition of the broadleaved deciduous forests. A vegetational survey of the Quercus aliena var. aculiserrata forests, which occupy an area of 6.800 m2, revealed a flora composed of 217 species belonging to 134 genera in 57 families, of which 126 species, or 58.1% of the total species, are endemic and 91 species are non-endemic. Most of non-endemic species are distributed throughout eastern Asia, and only a few species extend southward to Vietnam, Thailand and India, or to the Philippines and Australasia. The Quercus liaotungensis forests, occupying an area of 4,000 m2, are composed of 188 species in 118 genera and 53 families. Among these species 116, or 61.7% of the total, are endemic and 72 species are nonendemic. Of these non-endemic species about 54 are distributed throughout eastern Asia; the remainder extend south-ward to India, Bhutan, Nepal or Vietnam, and a few species reach the Philippines, Indonesia (Cayratia ja-ponica (Thunberg) Gagnepain) or Thailand (Disporum cantoniense (Lour.) Merrill).
In a forest of Betula albo-sinensis, occupying an area of 4000 m2, were 152 species, of which 111 of them, or 73% of all species, were endemic. Of the 41 non-endemic species, 26 are restricted to eastern Asia. Except for a few species extending northward to the Russian Far East, the remainder occur in Burma, Bhutan, India, Sikkim and Nepal. A Betula utilis forest, occupying an area of 6400 m2 was composed of 173 species, of which 119, or 68.8% of the total, were endemic and 54 were nonendemic. Among these non-endemic species 33 have their distribution in eastern Asia, 14 occur in Nepal, Bhutan, Sikkim, Burma and Vietnam. Only a few species, such as Hemiphragma heterophyllum Wallich are distributed southward to the Philippines. Another seven species are widespread in Eurasia and America.
2. Subalpine Coniferous Forests. An Abies fargesii forest with an area of 8400 m2 included 105 species belonging to 75 genera and 36 families. Among these. 72 species, or 68.6%, were endemics, including seven endemic to the Qinling mountains. Thirty three nonendemic species have distributions throughout eastern Asia; only six species extend southward to Vietnam. Burma, Thailand or Bhutan and India; three were cosmopolitan. Larix chinensis forests occur only in the Qinling Mountains. A survey of one such forest with an area of 6400 m2, revealed this community to be composed of 114 species, of which 76 species, or 66.7% of all species, were endemic and 38 species were non-endemic. Among the non-endemic species, nine have distributions in eastern Asia, ten occur in the north temperate zone, 11 extend southward to the mountainous regions of Burma, India, Bhutan, Sikkim or Nepal. The remainder are cosmopolitan or extend northward through Mongolia to central Asia.
Conclusions
Through an analysis of the composition of the various types of forests in the Qinling mountain range, it is clear that the vegetation can be thought of as comprising two main elements: an endemic or native ele-ment, and a Sino-Japanese element derived from the Tertiary Geoflora. The remaining small number of species are from miscellaneous sources.
Literature cited
- Boufford, D. E. and S. A. Spongberg. 1983.
- Eastern Asian-Eastern North American phytogeographical relationships-a history from the lime of Linnaeus to the twentieth century. Ann. Missouri Bot. Card. 70: 423-439.
- Chen, M. R. 1992.
- Regionalization of vertical temperature in Qinling mountain range. Geogr. Res. 11(3): 27-31.
- Cheng, M. 1984.
- The floristic relationship between eastern China and Japan. Acta Phytotax. Sin. 22: 1-5.
- Dansereau. P. 1951.
- Description and recording of vegetation upon a structural basis. Ecology 32: 172-229.
- Daubenmire, R. 1978.
- Plant Geography: with Special Reference to North America. Academic Press. New York.
- Graham, A.. ed. 1972 a.
- Floristics and Paleofloristics of Asia and Eastern North America. Elsevier Publ. Amsterdam.
- Graham, A. 1972 b.
- Outline of the origin and historical recognition of floristic affinities between Asia and eastern North America. pp. 1-18. In A. Graham, ed. Florislics and Paleofloristics of Asia and Eastern North America. Elsevier Publ. Amsterdam.
- Gray, A. 1846.
- Analogy between the flora of Japan and that of the United States- Amer. J. Sci. 2: 135-136.
- Hara, H. 1952-1956.
- Contributions to the study of variations in the Japanese plants closely related to those of Europe or North America.
- Hara, H. 1972.
- Corresponding taxa in North America, Japan and the Himalayas. pp. 61-72. In D. H. Valentine, editor. Taxonomy Phytogeography, and Evolution. Academic Press. London.
- Hu, H. H. 1935.
- A comparison of the ligneous flora of China and eastern North America. Bull. Chinese Bot. Soc. 1: 79-97.
- ———. 1936.
- The characteristics and affinities of Chinese flora. Bull. Chinese Bot. Soc. 2: 67-84.
- ——— and R. W. Chaney. 1940.
- A Miocene flora from Shantung Province, China. Contr. Paleobot. Carnegie Inst. Wash. Publ. 507: 1-147.
- Institute of Botany and Nanjing Institute of Paleontology, Academia Sinica. 1978.
- Cenozoic plants of China. Fossil Plants of China III. Science Press. Beijing. (in Chinese)
- Li. H. L. 1952.
- Floristic relationships between eastern Asia and eastern North America. Trans. Amer. Philos. Soc. 42:371-429.
- Lu, A. M. 1982.
- On the geographical distribution of the Juglandaceae. Acta Phytotax. Sin. 20:257-274.
- Mabberley, D. J. 1987.
- The Plant Book. Cambridge Univ. Press. Cambridge.
- Numata. M., editor. 1974.
- The Flora and Vegetation of Japan.Kodansha, Ltd. Tokyo.
- Shaw, H. K. Airy, revised. 1973.
- J. C. Willis's A Dictionary of the Flowering Plants and Ferns, 8th ed., Cambridge Univ. Press, Cambridge.
- Tanai, T. 1972.
- Tertiary history of vegetation in Japan. pp. 235-255. In A. Graham, ed. Floristics and Paleofloristics of Asia and Eastern North America. Elsevier Publ. Amsterdam.
- Wu, C. Y. 1979.
- The regionalization of Chinese flora. Acta Bot. Yunnan. 1: 1-22.
- ———. 1983.
- On the significance of Pacific intercontinental discontinuity. Ann. Missouri Bot. Card. 70: 577-590.
- ———. 1991.
- The Areal-Types of Chinese genera of seed plants. Acta Bot. Yunnan. Supp. 3: 1-139.
- ———. ed. 1980.
- The Vegetation of China. Science Press. Beijing. (in Chinese)
- Ying, T. S. 1983.
- The relationships of the temperate forest regions of China and the United States. Ann. Missouri Bot. Gard. 70: 597-694.
- ———, Y. F. Li, Q. F. Guo and H. Cui. 1990.
- Observations on the flora and vegetation Taibai Shan, Qinling mountain range, southern Shaanxi, China. Acta Phytotax. Sin. 8: 261-293.
- ———, S. Terabayashi and D. E. Boufford. 1984.
- A monograph of Diphylleia (Berberidaceae). J. Arnold Arbor. 65: 57-94.