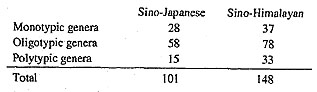Delineation and Unique Features of the Sino-Japanese Floristic Region
Cheng-yih Wu
Kunming Institute of Botany, Chinese Academy of Sciences, Kunmimg 650204, Yunnan, China
|
Abstract. Based on the definitions of the Eastern Asiatic Region by Diels and sensu Takhtajan, and the Sino-Japanese Region sensu Good, the historic changes in the delineation and subdivision of this region are concisely discussed. The separation of the Sino-Himalayan Subregion and the Sino-Japanese Subregion (in a strict sense) is proposed, with the former including provinces 9-13 sensu Takhtajan. The East China Province and South China Province are separated from several neighboring provinces, such as the Indo-Malayan Province of the Paleotropics in the broad sense. The diversification of both new subregions and provinces, with a brief statement on the unique characteristics of the whole region, are also summarized on the basis of an analysis of the genera of the region. The floristic interrelationships among these subdivisions are also briefly discussed. Key words. Sino-Japanese Floristic Region, Eastern Asiatic Floristic Region, phytogeography, eastern Asia The Sino-Japanese Floristic Region (essentially the same as the East Asian Region of Diels, 1901; East Asiatic Floristic Region of Drude, 1890, and Temperate Eastern Region of Engler, 1912) has been recognized as a natural floristic area since Grisebach first discussed it in 1872. The delineation and subdivisions (regionalization) of the Sino-Japanese Floristic Region, however, is still a problem open to discussion and argument. In his book. The Floristic Regions of the World, Takhtajan (1978, p. 50; 1986, p. 39) very concisely summarizes the history of the region and defines its boundaries. We agree with his view in general, but prefer to use more natural boundaries. In northeast Asia the Sino-Japanese Floristic Region is bounded by the Siberian Taiga and the northeastern Korean pine-mixed forest, to the northwest the boundary is the transition zone between forest and steppe or subalpine scrub meadow. To the southwest and south the natural boundary is the evergreen pine-oak forest and the mixed evergreen-deciduous monsoon and rain forest, with the former being dominated by Fagaceae, Lauraceae, Hamamelidaceae and Magnoliaceae, but mixed with various conifers and taxads, especially Pinus. The latter forests are dominated by the large pinnateleaved families (Fabaceae in the broad sense, Meliaceae, Sapindaceae, Anacardiaceae, Burseraceae, etc., with remarkably many species of Ficus), or by Dipterocarpaceae, Tetramelaceae, Crypteroniaceae, etc. These forest have few intermixed conifers, but contain a few cycads and podocarps. The floras of this vast region are extremely rich, diverse and unique, and contain no fewer than 30 endemic families, or more than two times the number of endemic families recognized for the region by Takhtajan in 1978 (Table 1).
By analyzing this region the following observations can be made: 1) the area is characterized by the presence of very old lineages of gymnosperms; 2) most of the gymnosperms are monotypic (16) or oligotypic (11), and only 2 are polytypic-this phenomenon certainly denotes antiquity and the generally relict nature of the region; 3) the dispersion among several important evolutionary stocks, especially in the "polypetalae" and apetalae," and again centering in the Magnoliidae, Ranunculidae and Hamamelidae sensu Takhtajan; 4) the scarcity of endemic families in "sympetalae" and "monocotyledoneae," with only three in the former and only one in the latter; 5) the mostly woody nature of the families (20 families, or almost two thirds of the total, including two woody lianas). These facts are certainly strong support for the hypothesis that the birthplace and cradle of the Hoiarclic flora may be here. In addition, about eight families extending to tropical Southeast Asia (Indo-Malaya) are also noteworthy. They are Nageiaceae (1/6-one genus/6 species), Rhodoleiaceae (1/1 (7-10)). Daphniphyllaceae (1/9-10),Pentaphyllaceae (l/1).Duabangaceae (1/2), Mastixiaceae (2/15), Pentaphragmataceae (1/25) and Lowiaceae (1/7). This list shows approximately the same status of these families to the endemic families of eastern Asia and strongly suggests that the eastern Asian and southeast Asian floras are derived from the same ancestral stock. A comparison of these families with some of those mentioned above, such as the Plagiopteraceae, Triplostegiaceae and Carlemanniaceae, the traces of the floristic evolution and floristic interrelationship among such floras, that is, the eastem Asian flora, the western Malesian flora and the Indo-Malayan flora, are also quite obvious. Takhtajan (1978,1986) estimated that there are more than 300 endemic genera in his Eastern Asiatic Region, including pteridophytes. To date, the endemic genera that we have enumerated from the botanical provinces IE 10-18, 11G 19, on our map of the botanical regionalization of China (C. Y. Wu and S. K. Wu, 1993) already numbers 214, a majority of the endemic genera of the Eastern Asiatic Region. This subregion includes only eastern China east of a line that passes from northeast China to southwest China. If the genera endemic to the entire Eastern Asian Region (71) (Table 2), the genera endemic to the Sino-Himalayan Subregion (148) (Table 3), the genera endemic to the Sino-Japanese Subregion (101) (Table 4), and the genera endemic to Japan, Korea, the Bonin Islands and the Ryukyu Islands (42) (Table 5) are included the total number of endemic genera in the whole East Asiatic Region is estimated to be more than 577 genera (excluding pteridophytes). These facts strengthen support for the words of Takhtajan (1978, 1986:48). Thus the Eastern Asiatic Flora is characterized by an exceptionally large number of endemic genera, which are in large part monotypic or oligotypic. These genera belong to very different families, both archaic and advanced. Yet many of these endemic genera belong to the archaic subclasses Magnoliidae, Ranunculidae, and the Hamamelididae. This underscores the antiquity of the flora, which contains not only many ancient relict endemics of various taxonomic ranks, but also a series of endemic families and even suborders.
The flora of the Eastern Asiatic Region holds special interest for the study of the history of the temperate floras of the northern hemisphere, as well as for the solution of many problems of evolution and systematics of the flowering plants. The Eastern Asiatic Region appears simultaneously as one of the major centers of development of higher plants (especially the gymnosperms and angiosperms), and as one of the centers of the preservation of ancient forms (gigantic refuges of "living fossils"). Here one finds many archaic and intermediate forms that are of great importance in the elucidation of phylogenetic problems. Many genera, and even families, that are well differentiated in the flora of the Circumboreal Region are here connected by intermediate taxa, and often merge. Now we will discuss the subdivisions of this huge flora. The separation of the Sino-Malayan subregion and the Sino-Japanese subregion (in the strict sense) is necessarily proposed here with the former including provinces 10-13 and the latter provinces 1-9 of Takhtajan (1986). First of all, we should combine both the floristic and vegetational characteristics. The differentiation of this separation is supported by many features, yet there is also a homogeneity shown by the endemism in the whole Eastern Asiatic Region as represented by almost 71 genera in 45 families of the Eastern Asian distribution pattern that are all present in mainland China. They are variously associated in diversified ecosystems, with various distribution patterns overlain on others, such as the eastern Asian-eastern North American distribution pattern overlaying the whole North Temperate pattern. These distribution patterns are variously associated into diversified ecosystems from Northeast China (mixed deciduous forest with Pinus koraiensis as one of the dominants). North China (drier mixed deciduous oak and pine forests). East China (mixed deciduous and broadleaved evergreen forest, interspersed with large patches of Pinus massoniana, Cunninghamia and Cryptomeria), Central China (same as East China, but with more deciduous woody endemics and various conifers and taxads). South China (Southeast China Province of Takhtajan, 1986) (mainly evergreen pine-oak forests, but with many tropical elements from the Indo-Malayan region) to Yunnan-Guangxi-Guizhou (mainly limestone vegetation of mixed forests with very diverse elements common to the Gulf of Tonkin and the adjacent Yunnan-Guizhou Plateau intermixed. These six provinces together with the Taiwanian Province in the sense of Takhtajan (IIG 19 in the sense of C. Y. Wu and S. K. Wu) are dominated by genera and species endemic to the Sino-Japanese region. From IF 16-18, these three provinces, in contrast, are dominated by genera and species characteristic of the Sino-Himalayan region and forming various forest types and scrub and meadow systems somewhat reminiscent of those in the Mediterranean region. Successively from south to north and northwest on the western edge of the Sino-Japanese Region are the Yunnan Plateau (evergreen and deciduous oak forests, or sclerophyllous oak forests intermixed with Pinus yunnanensis, or rain shadow effect savanna-or marquis-like vegetation in deep valleys), Hengduan Mountains (vertical distribution of forest to subglacial ecosystems starting from dry, riparian vegetation up to and dominated by subalpine coniferous forest resembling mountain taiga) and the eastern Himalaya-North wing (same as Hengduan Mountains, but impoverished from east to west in endemism in both genera and families). The last province is merely an extension of 12. The Eastern Himalayan Province, which is closely related to the Northern Burmese Province (11) and the Khasi-Manipur Province (13) in the sense of Takhtajan (1986) is also dominated by Sino-Himalayan genera. The Flora of Japan, the Bonin and Ryukyu islands, and KoreaI would first like to discuss the floras of Japan, the Bonin and Ryukyu islands and Korea. The area from northeastern Japan (Hokkaido) to Kyushu, the Bonin and Ryukyu islands and from North to South Korea epitomizes the eastern part of mainland China. Although this area contains only 42 endemic genera (as far as I am able to determine), comprising less than 20 percent of the total number of endemic genera of the forested parts of southeastern China, it is very similar to the latter in that it contains three monotypic, endemic families; namely the Glaucidiaceae, Pteridophyllaceae and Sciadopityaceae. The former two are perennial herbs that are closely related to the rather primitive families of Ranalian affinity, the Podophyllaceae and Papaveraceae, respectively. Sciadopitys, an isolated member of woody, tree-sized gymnosperms, is related to the relatively primitive Taxodiaceae in the Coniferales. These three families, two herbaceous and one woody, are certainly relicts and epibiotics. In China, however, the three (or two) monotypic endemic families are all woody, primitive and epibiotic. Oligotypic genera in this area total eight, or about one quarter the number of monotypic genera. Their life forms include one tree, three shrubs and four herbs. In addition to Asteraceae and Orchidaceae, they occur only in the Rutaceae. Thymelaeaceae, Diapensiaceae, and Campanulaceae. The remaining 30 or so genera are monotypic, and except for the three families mentioned above, belong to the Orchidaceae (7 genera), Asteraceae, Ranunculaceae (each with 3 genera), Apiaceae, Ericaceae, Lamiaceae, Melanthiaceae (Liliaceae), Rosaceae, Scrophulariaceae (each with 2 genera), Araliaceae, Campanulaceae, Cupressaceae, Podophyllaceae, Euphorbiaceae, Urticaceae, Oleaceae, Boraginaceae, Liliaceae, Anthericaceae (Liliaceae), Arecaceae, Poaceae, Thymelaeaceae (each with 1 genus). Six genera, including Abeliophyllum (Oleaceae), Diplolabellum (Orchidaceae), Hanabusaya (Campanulaceae), Megaleranthis (Ranunculaceae) and Pentactina (Rosaceae) occur in Korea, Boninia (Rutaceae), Boninofatsia (Araliaceae) and Dendrocacalia (Asteraceae) are in the Bonin Islands, while Satakentia (Arecaceae) is only in the Ryukyu Islands and on Sakishima Island. In this incomplete list both Thujopsis (Cupressaceae) and Satakentia are noteworthy from a phytogeographical point of view. In short, it is reasonable to keep the original Sakhalin (Kuriles) Hokkaido Province (Tolmachev, 1899), the Japan-Korean Province (Good). the Volcanic-Bonin Province (Engler) and the Ryukyu Province (Tokara-Okinawa Province) (Engler) as separable and distinct in a floristic sense, but they should be classified under the Sine-Japanese Subregion in our point of view. In mainland China there are perhaps 101 genera in 53 families belonging to the Sino-Japanese distribu tion pattern, while 148 genera in 52 families belong to the Sino-Himalayan distribution pattern. For comparison we have arranged the families of the two areas by number of endemic genera in Table 6.
Table 6 reveals some surprising dissimilarities in the two areas, even though they are both temperate in nature. They differ in the following:
From the observation outlined above, 1 feel that the separation of the whole Eastern Asiatic Region into two subregions, a Sino-Japanese subregion and a Sino-Himalayan subregion, is more in line with the objectives of florogenesis and more clearly indicates the distinctiveness in their floristic composition. Other unique characteristics of the Eastern Asiatic Province on the Chinese mainlandIE 10. Northeastern China Province (=Manchurian Province, Krasnov, 1899). Since Krasnov (1899), Good (1964) and Takhtajan (1978, 1986), this province is well known, but Transbaicalia and Eastern and Northeastern Mongolia should be removed from the forested Eastern Asiatic Flora and treated as part of the Eurasian Steppe. Although mixed deciduous forests marked by Pinus koraiensis cover vast mountainous regions and contain a number of old floristic elements, endemic genera are very scarce and no endemic families occur within this province. Six (or seven) herbaceous genera are characteristics of this flora (Table 8), but none are strictly endemic to China. Two of them belong to the primitive Saxifragaceae, and one, Plaeiorhegma, which is a counterpart (or congeneric) of the North American Jeffersonia, to the even more ancient family Podophyllaceae, while four of them extend to North Korea. Brachybotrys (Boraginaceae), which occurs in forests, and Symphyllocarpus (Asteraceae), an inhabitant of swamps, are both endemic to northeast Asia in general. Only one saprophytic orchid with rather primitive characters also has been recognized recently for its relict, epibiotic nature. Microbiota, sometimes reduced to Thuja, is not found on the Chinese side of this province, while Omphalotrix (Scrophulariaceae) is classified by us also as a characteristic genus of the Eastern Mongolian, Central Manchurian and Eastern Siberian Steppe.
IE 11. Northern China Province. This is the seventh province in Takhtajan's scheme. However the northwestern limits, which were adopted by Takhtajan from us, lie on the demarcation line between the Bothriochloa meadow together with "aestisilvae" and the Stipa-Artemisia steppe. The characteristic vegetation here is a drier mixed deciduous oak forest marked by Pinus tabulaeformis or P. densiflora (from maritime areas to the Liaotung and Shandong peninsulas). About 16 or 17 endemic (6 strictly endemic) or characteristic genera, both monotypic (10) and oligotypic (7), are present here (Table 9). Two of them, one in Ulmaceae (Pteroceltis, which extends to central, east and southwest China) and one in Sapindaceae (Xanthoceras) attain tree size. Apparently all are relict and epibiotic. Among the four shrubs, one is in the rather ancient family Corylaceae, one in the Rosaceae, one in Caprifoliaceae, and one in the woody primitive tribe Mutisieae in Asteraceae. There are two herbaceous vines, one in Polygonaceae and one in Cucurbitaceae. It is interesting to note the distribution pattern of the bispecific Bolbosfemna, one species ranges from northeast to central China, while the other occurs disjunctly on the Yunnan Plateau. There is also a small disjunction, represented by two varieties, between Jinshajiang and the Red River. This pattern mirrors the one in the shrubby Ostryopsis (Corylaceae). in which there are also two closely related species, one found only in Jinshajiang and the other in north China. This pattern strongly suggests the Gondwanan origin of these tertiary relictual and epibiotic genera. Sinoplagiospermum also displays a dispersed distribution, seeming closely related to the Sino-Himalayan Prinsepia, and is probably also relictual and epibiotic. We can probably safely conclude that north China has a rather ancient flora that dates from the Tertiary.
IE 12. East China Province. This is the east wing of the "Central China" province sensu Diels (1901). It lies on the "first staircase" of Chinese topography, and is probably the native home of Ginkgo, Pseudolarix and Pseudotaxus. Of the 22 endemic and characteristic genera in this province, at least 9 are strictly endemic to the eastern lowlands and hills (Table 10). Cunninghamia seems to have been native to this area and is now propagated in the whole region south of the Chang Jiang (Yangtze) and north to the Qinling Mountains and Huai-he. Ginkgo, in a semi-wild state at Tianmu-shan (Zhejiang), is most noticeable as a characteristic monotypic relict family here. Twenty one genera in 18 families include 7 genera of trees, 5 of shrubs, 1 woody liana, and 8 genera of herbs. One of the two woody hamamelid representatives, Shaniodendron, is very interesting because of its recent discovery as a member of a tribe formerly not recorded in China or in Eastern Asia. Sinocalycanthus (Calycanthaceae) may also indicate the antiquity of the Eastern Chinese Flora. Even in Brassicaceae. Lamiaceae and Orchidaceae, we have recently recorded such genera as Hilliella, Cochleariella, Sinopogonanthera and Nothodoritis. The paucity of species in the 21 genera (17 are monotypic, 3 are oligotypic and only 1 is polytypic) characteristic of this province may be also an indication of its antiquity.
IE 13. Central China Province. This is certainly the west wing of the former "Central China" Province of Diels (1901), sensu stricto, and seems to be the most typical and condensed one as a representative for the whole Chinese Flora. Although Central China, like Eastern China, was in prehistoric times densely clad with evergreen oak forests with many lakes and swamps, only fragments of the original forest still exist. This region is marked by Pinus massoniana and other thermophilous conifers and taxads at low elevations and gradually is transformed northward and upward into mixed deciduous and evergreen oak forests or beech forests with many woody genera and families of the eastern Asiatic pattern, Sino-Japanese pattern and eastern Asia-North American pattern. The differentiation of the East and West still seems very striking. Among the 51 genera mentioned (about 27 are strictly endemic), with the exception of only 4 neoendemic polytypic genera (Ancylosiemon and Jspmeimm (Gesneriaceae), Qiongshuea (Bambusaceae-Poaceae) and Notoseris (Asteraceae)), nearly all genera are monotypic (30) or oligotypic (17), and probably no less than 30 are relict and epibiotic (Table 11). Like Ginkgo in Eastern China, the famous genera are Cathaya (Pinaceae), Meiasequoia (Taxodiaceae), Eucoinmia (Eucommiaceae), Bretschneidera (Bretschneideraceae) and Davidia (Davidiaceae). The latter 3 are also characteristic endemic monotypic families for the whole of China, although Bretschneideraceae has been found recently also in northern Laos to northern Thailand and northern Vietnam. Among the 51 genera already mentioned, 11 are trees, 7 are shrubs, 30 are lianas and 26 are herbs. In addition to the 5 genera just mentioned, Camptotheca (Nyssaceae), Chimonanthus (Caly canthaceae), Dipteronia (Aceraceae), Poliothyrsis (Flacourtiaceae), Poncirus (Rutaceae), Sinojackia (Styracaceae), Sinowilsonia (Hamamelidaceae) and Tetrapanax (Araliaceae) are trees from some relatively primitive families or occupy a primitive position within their family. All 3 woody lianas, Clematoclelhra (Actinidiaceae), Sargentodoxa (Sargentodoxaceae) and Sinofranchetia (Lardizabalaceae) belong to primitive famities. Sargentodoxa represents an endemic monotypic family extending from northern Vietnam northward to the Qinling Mountains. Saruma (Aristolochiaceae), Eomecon (Papaveraceae), Psilopeganum (Rutaceae) and Dickinsia (Apiaceae) are herbaceous representatives of this pattern that occupy a somewhat more primitive position within those primitive families of the Magnolidae, Ranunculidae and Rosidae. Recently, as in the East China Province, several herbaceous endemics were discovered from this province.
IE 14. South China Province. The South China Province is segregated from Kingdom II, Paleotropic Subkingdom, Indo-Malaya, Indo-China region as a part of the South China Province of Fedorov (1958). It occupies a narrower region, mainly south of the Nanling Mountain Range, which extends across the northern borders of Guangxi and Guangdong to Fujian (the Peiling is represented by the Qinling Mountain Range), and north of the Leizhou Peninsula in Guangdong Province, and includes a part of southeast Guangxi. The main vegetation cover here is a transitional evergreen pine-oak forest but with several Indo Malayan elements represented by trees, undershrubs, lianas and herbs. The southern boundary lies on the northern limit of such typical tropical families as Dipterocarpaceae, Myristicaceae, Nepenthaceae, Ancistro cladaceae and Flagellariaceae. About 25 genera (14 strictly endemic) may be considered as endemic or characteristic of this province (Table 12). Seven genera attain tree size, 3 are shrubby, 2 are lianas and 13 are herbs. With the exception of the polytypic Indocalamus, a shrubby bamboo, all the genera are either monotypic (18, about 2/3 of the total) or oligotypic (8, or nearly 1/3). Two genera of Hamamelidaceae, Semiliquidainbar in the Nanling Range, and Tetrathyrium in Hong Kong, and two theaceous genera, Apterosperma and Euryodendron, strongly suggest the Holarctic-Eastern Asian nature of the South China Flora, yet the presence of Sinia (of Sauvagesiaceae, a segregate of the pantropical Ochnaceae) provides a link to Indochina and Indonesia. Two melastomataceous genera, two rubiaceous genera and five gesneri aceous endemic genera show the strong influence from the Indo-Malayan Subkingdom of the Paleotropics. Glyptostrobus (Taxodiaceae) is a characteristic remnant of the maritime lowland plain of South China from ancient times. It also represents a generic counterpart of Taxodium of the southeastern United States and northeastern Mexico in North America. There are nearly five genera characterizing the Nanling region, among which Nothotsuga is a primitive newly established genus in the Pinaceae, and may be a living link between Tsuga, Pseudotsuga. Cathaya. and Picea. Considering the incidence of endemism and the richness of woody plants, the South China flora is comparatively less primitive than the floras of the Eastern and Central Chinese provinces, although it certainly belongs to the Eastern Asiatic Region rather than to the Indo-Malayan Subkingdom.
IE 15. Yunnan-Guangxi-Guizlioii Province. This floristic province was little known at the time of Takhtajan (1978, 1986). It perhaps belongs to parts of his Central China and South China provinces. We now delineate this Province as comprising southeast Yunnan (the Wenshan region above 1000 m), Guangxi (most parts of the Hongshui He and Youjiang regions), and southwest and southern Guizhou (Libo, Dushan and Loudian west to Xinyi). This Province presents a very rich and relatively ancient limestone flora dominated by southern subtropical evergreen oak forests (many endemic and primitive "Cyclobalanopsis," Castanopsis and Lilhocarpus) together with Keteleeria and a few limestone-loving 5-needled pines. The high percentage of monotypic genera is remarkable. Of the 29 genera (21 strictly endemic) listed in Table 13,23 are monotypic and 4 others are oligotypic. Although there are no endemic families, the Rhoipteleaceae, classified in the Tonkin Bay Flora of the Indo-China Region, extends to southern Guizhou and southeast Yunnan, and Dipentodontaceae is frequently met with here. Five trees, 4 shrubs, 1 palm, 2 woody lianas and 17 herbs make up the characteristic genera of this flora. Among them several genera are remarkable, including one in the Magnoliaceae (Manglietiastrum), three in Sapindaceae (Delavaya linked with the Yunnan Plateau, Eurvcorymbus linked with Central China and Taiwan, and Handeliodendron linked with South China) and one in the Olacaceae (Malania). They suggest the tropical origin of some Holarctic Eastem Asian elements, but not from Indo-Malaya. Other genera arc concentrated in Gesneriaceae (up to 14) and Bambusaceae (2) with Apocynaceae, Asclepiadaceae, Melastomataceae and Orchidaceae (each 1). These taxa seem to denote that although the transitional position of this floristic province is obvious, it stillbelongs to Eastern Asia as a southern subtropical limestone variety. The recently discovered Saniculiphyllum in the Saxifragaceae and the palm genus Guihaia are also very interesting from this florogenetic point of view.
IF. Sino-Himalayan Forest Flora Region. The unique characteristics of this region were presented in my paper on the regionalization of the Chinese flora (Wu, 1979), which reads as follows. This is another very rich and ancient flora distributed between 20-40 N. Lat. from temperate region through the subtropics to the northern boundary of the tropics. It is suspected to include the Himalayan Region outside China. The total number of species may be over 20,000 (the Yunnan Plateau and part of the Hengduan Mountains aione are estimated to contain over 12,000 species). On one hand, owing to the complicated topography and climate, many natural refugia have resulted in the preservation of many relict plants from the Tertiary or even before; but on the other hand, owing to the great altitudinal changes, the rapid and continuous uplifting and creation of new habitats, many neotypes have appeared successively causing the preservation of intermediate stages during the evolutionary process. The floristic elements, therefore, are a combination ofepibiontics and neoendemics, being a mixture of elements from all corners of the earth, with very obvious altitudinal distribution, and sometimes combined in their horizontal distribution to form different zonational bands at different altitudes, and sometimes concentrating many (upwards to 8) vertical zones within a very short distance. The unique and genera] characteristics are that the forest vegetation at lower altitudes is similar to that in the Sino-Japanese region (IE), also being composed of the same generic series, but the species of thermophilous conifers and taxads and the dominant "Cyclobalanopsis" Castanopsis and Litlwcarpus are quite different and are vicarious species of the former region. The successive appearance of Tsuga, Picea, Abies, Larix and Juniperus (Sabina) in the subalpine and alpine zones is also in the same order as they appear in the alpine zone in central China, but a series of vicariants is also present and often forms a broader horizontal zonation in combination with the altitudinal distribution. Calocedrus and Keteleeria are substitutes for Chamaecyparis (Taiwan) and Pseudolarix in IE. In addition to the special richness of woody plants and the ancient and relict families, genera and species, the trees, shrubs, lianas and herbs of the alpine and subalpine zones are also specially developed. IF 16. Yunnan Plateau Province. The Yunnan Plateau Province was included in the Sikang-Yunnan Province (9) sensu Takhtajan (1978), but we separate out the northern part of the higher altitude area north ward from Lijiang and the Zhongdian Plateau into the Hengduan Mt. Province (IE 17) below. The Yunnan Plateau Province lies on the Sikang-Yunnan ancient landmass and enjoys a much wanner and drier climate in comparison with the Hengduan mountain Province. All 19 genera endemic or characteristic of this province (Table 14) are monotypic (16) or oligotypic (3. only about 1/6 of the total). Except for 1 tree, 1 small shrub and 1 bamboo, the remainder are herbaceous or essentially so. The tree genus Dichotomanthes (Ro-saceae) is very isolated and notable for its systematic position and primitiveness, as are also the primitive labiates Cardioteucris and Holockeila, which link the Lamiaceae and Verbenaceae. The specialized but dispersed genus Psammosilene (Caryophyllaceae) and Bolbostemma (Cucurbitaceae) (in list of genera IE 11) certainly reveal the complicated nature of their origin and dispersal. But Rhabdothamnopsis (Gesneri-aceae) and Musella (Musaceae) obviously show their relationship with African relatives. Overall, this flora is rather old and complicated, with certain relationships both to the Laurasian, Gondwanan and Tethyan elements.
IF 17. Hengduan Mountain Province. The Hengduan Mountain Province occupies the bulk of the former Sikang-Yunnan Province of Takhtajan, but does not extend southward below the Lijiang and Zhongdian Plateau. It extends to the Taro River drainage area in Gansu up to its Qinghai boundary, and is thus connected to Tangut Province of the Qinghai-Xizang Plateau Floristic Region. The delineation and unique characteristics of this province are addressed in two recent papers by Wu (1988) and Li (1993) and are not discussed further here. Among the 38 genera listed in Table 15, 27 appear to be strictly endemic to this province.
IF 18. Eastern Himalaya North Wing Province. This province is essentially the Eastern Himalaya Province of Hooker (1904). but not including the tropical pans of the southern slopes of the Himalaya, and mainly the subalpine coniferous forest region in southeast Xizang around Nantschabawa, extending to the Himalayan countries from Bhutan and Sikkim to eastern Nepal. This province is covered for the most part by successive zones beginning with Pinus yunnanensis, P. densata, P. griffithii and Quercus semecarpifo-lia on the southern drier slopes and other evergreen oaks in deep valley. This is an impoverished variety of the Hengduan Mountain Province, but with several (up to 8) zones on the southern slopes. There is no generic or familial endemism developed, but strong vicariance phytogeography can be observed. Recently, only two herbaceous genera, the monotypic Xizangia and the oligotypic Sinoleontopodium, have been added to this province on the Chinese side (Table 16). They are certainly neoendemics that evolved after the upheaval of Nantschabawa and are probably derived from the neighboring genera Phtheirospermum and Leontopodium respectively. Perhaps Bryocarpum, Cavea, Milula and Platystemma, with a distribution from the eastern Himalaya to Xizang, should be listed here also. There are perhaps another 14 genera endemic to this province that have not yet been recorded on the Chinese side.
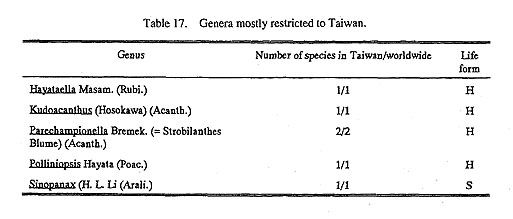
Other provinces suggested by Takhtajan (1986) such as the Northern Burmese (now Myanmar) Province, and Khasi-Manipur Province, owing to a lack of a concrete knowledge of the flora, are tentatively omitted here. The Taiwanian Province of Takhtajan (1986) is also removed from here to the Paleotropic Kingdom because of the floristic features of the lowlands and especially of the southern peninsula and southern neighboring islands of Lanyu and Lutao. Although the northern half of the island, especially the higher mountains, supports a rich flora and vegetation parallel to that of the Himalaya yet: 1) there are elements, even in the alpine zone, from the southern hemisphere (for example, Oreomyrrhis); 2) the few strictly en-demic-or nearly endemic genera (Table 17) are mostly from the lower altitudes; 3) the general floristic com position, even northward to Taibei, includes many tropical families and genera similar to those of Hainan Island; 4) there are well developed mangrove forests, even including Nypa palm and Suriana (Surianaceae), and the well-developed strand forest with Barringtonia, Guettarda, Hernandia, Heritiera, Pisonia, Pem-phis and others. From this evidence, I prefer to treat Taiwan as a whole as belonging to the northernmost part of the Paleotropics, like IIG 23 Eastern Himalaya South Wing in our scheme.
|