Pronucula insignis sp. nov.
Figures 16-25
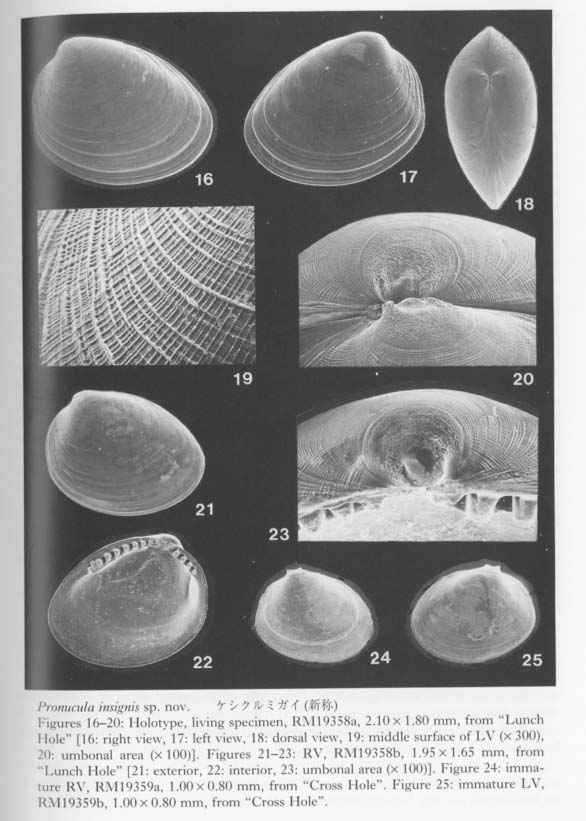
Type and material.—Holotype: RM19358a, a living specimen from the bottom sediments of "Lunch Hole" of Irabu Islet, Miyako Islands. Paratypes: RM19358 (living) from the type locality, RM19356 (dead) from "Devil's Palace", RM19357 (dead) from "Toriike", RM19359 (dead) from "Cross Hole" of Shimoji-Irabu Islet. RM19360 (dead) from "Shodokutsu" of Ie Islet.
Diagnosis.—Small subtriangular species of Pronucula, characterized by truncated posterior margin, subquadrate resilifer, relatively few paleotaxodont teeth, numerous divergent threads on external surface, finely crenulate inner margin, and large saucershaped Pd I with an irregularly depressed summit.
Description.—Shell rarely exceeds 2.2 mm in length, nacreous, translucent, subtriangular, about 1.3 times longer than high. Periostracum pale brownish, very thin. Antero-dorsal and postero-dorsal margins continuously arcuate below umbo, but forming obtuse angles with anterior and posterior margins; the former much longer than the latter. Umbo orthogyrous, located about a third of a shell length from the front. Surface nearly smooth except for numerous faint divergent threads, which almost perpendicularly cross irregular growth lamellae. Inner margin of valve very finely crenulate in accordance with subinternal radial threads only in adult stage. Resilifer deep and large, subquadrate, situated just below umbo, flanked by a pair of symmetrical tooth-like projections, not forming an oblique chondrophore. Teeth tubercular, about eight on anterior and three or four on posterior hinge plate in the largest specimens. Pd I very large, 240-285 µm in maximum diameter, saucer-shaped, with irregularly depressed summit and pockmarked surface; Pd II absent.
Remarks.—This is a solitary paleotaxodont species from these submarine caves. Several individuals from the type locality ("Lunch Hole") are considered to be adult. All the dead specimens (e. g. Figures 24, 25) from other caves of Shimoji, Irabu and Ie Islets, however, are much smaller (rarely exceeding 1.2mm) and have fewer paleotaxodont teeth than those from the type locality, though the prodissoconch and surface features are the same. It is unknown whether these smaller specimens indicate a stunting as an adaptation to the cavernicolous environment or represent only the juvenile stage.
The huge saucer-shaped Pd I (Figures 20, 23) seems to characterize the present species because such a feature has scarcely been described in nuculids. The posteriorly truncated outline and subinternal radial threads may remind one of those of Nucula paulula Adams, 1856, a common Japanese nuculid, but, so far as we observed the specimens of that species from several localities, the shell grows much larger, and the Pd I is much smaller and not saucer-shaped.
In view of the undeveloped chondrophore, continuously arcuate dorsal margin and relatively few palaeotaxodont teeth, this species is referable to Pronucula rather than Nucula (s. s.). It resembles Promicula decorosa Hedley, 1902, the type species of this genus, from the lower sublittoral bottom of New South Wales, but the outline is more trigonal and the posterior part is more reduced. In the truncated posterior margin it resembles Nucula minuta Tenison-Woods, 1878, from the same region, which was also referred to Pronucula by Hedley (1902), but its radial (or divergent) threads may be weaker. Pronucula maoria Powell, 1937, if his illustrated specimen is adult, has fewer anterior teeth. Nucula certisina Finlay, 1927, the holotype of which was refigured by Dell (1956), reveals similar hinge structure to the present cave species, but the shell is taller and probably the external surface is covered with more prominent growth lamellae. According to Powell (1958) and others, some species of Austronucula from Australia and New Zealand reveal somewhat similar outlines to the present species, but the genus is generally characterized by entire inner margin.
LaBarbera (1974) examined the larval and post-larval shell development of an incubated species, Nucula proxima, from the Miocene of Florida. In that species prereleased juveniles (smaller than 600 µm in length) show quite different morphology from the adults, e.g. subquadrate resilifer flanked by a pair of symmetrical tooth-like projections, a few tubercular palaeotaxodont teeth and numerous fine diverging threads, all of which are shared with the present new species. It is, therefore, likely that the present and other species of Pronucula were derived from Nucula species through paedomorphic evolution.
Distribution.—This species has been found in a few sublittoral caves of Shimoji, Irabu and Ie Islets, Ryukyu Islands.
Subclass Pteriomorphia Beurlen, 1944
Order Arcoida Stoliczka, 1871
Superfamily Arcoidea Lamarck, 1809
Family Arcidae Lamarck, 1809
Cryptic arcids from the submarine caves of the Ryukyu Islands have great species diversity; at least six diminutive species can be distinguished in the present samples. Only two of them, however, have been found alive, and most specimens are disarticulated valves.
Many of these cave arcids seem to be related and are regarded as epibyssate species in view of the subtrapezoidal shell and nearly straight (or slightly concave) ventral margin. They share a more or less reduced anterior part, a small number of prionodont teeth, sharply truncated posterior margins and well-developed commarginal lamellae on the external surface. These characters are indicative of a deep-water genus Bentharca. Yet their generic assignment is not very conclusive, because the posterior carina and radial ornaments are generally stronger than those of typical deep-water species of this genus, and because some of them are also similar to shallow-water species of Acar. Knudsen (1967, 1970) synonymized Bentharca with Acar, while Oliver and Allen (1980) and many others regarded them as distinct genera. As noted by Habe (1977a) and Oliver and Allen (1980), Bentharca has been represented by several Atlantic and northern Pacific species, the occurrences of which have been restricted to bathyal-abyssal depths, at least in low-middle latitudinal regions. Interestingly, these cavernicolous arcids, except one species, are regarded as non-planktotrophic, because of the very large Pd I and the absence of Pd II. This is unusual for shallow-water arcids in low latitudinal regions. Although the cardinal area commonly attains a certain breadth with growth, alivincular ligament sometimes persists until the adult stage, suggesting a paedomorphic feature. The alivincular ligament is situated just below the primary ligament of prodissoconch.