Systematic Treatments
L. Thunbergii (DC.) Nakai
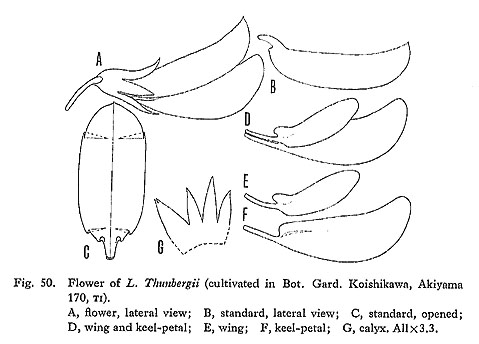
Lespedeza Thunbergii (DC.) NakaiLesp. Jap. Korea, 15 (1927), excl. syn. L. formosa (Vog.) Koehne-Ohwi, Fl. Jap. rev. ed., 790 (1965); Fl. Jap. Eng. ed., 559 (1965); Fl. Jap. new ed., 790 (1975), excl. var. obtusifolia (Nakai) Ohwi, var. albiflora (Schneid.) Ohwi, var. satsumensis (Nakai) Ohwi-Hatusima in Mem. Fac. Agr. Kagoshima Univ. 6: 11 (1967), excl. f. sericea Hatus.-Ohashi in Satake et al., Wild. Fl. Jap. Herb. 2: 205 (1982), excl. f. macrantha (Honda) Murata, var. satsumensis (Nakai) Ohwi. [Plates 26 & 27; Fig. 50]
Hedysarum heterocarpon (non L.) Thunb., Fl. Jap., 287 (1784). Desmodium Thunbergii DC., Prodr. 2: 337 (1825). D. racemosum (non DC.) Sieb. et Zucc. in Abh. Muench. Akad. 4. Abt. 2:121 (1845), pro parte. D. penduliflorum Oudem. in Neerlands Pl. 2: t. 2 (1866)-Carriere in Rev. Hort. 45:211, cum. tab. col. (1873). L. Sieboldii Miquel in Ann. Mus. Bot. Lugd.-Bat. 3: 47 (1867), excl. quoad specim. cit. Textor, Oldham 333 et Prol. Fl. Jap., 235 (1867), pro pane-Franchet & Savatier, Enum. Pl. Jap. 1: 101 (1875)-Bailey, Cyclop. Americ. Hort. 2: 904, fig. 1264 (1901)- Schneider, Ill. Handb. Laubholz. 2: 113, fig. 71e-g (1907)-Makino & Nemoto, Fl. Jap., 736 (1925), excl. var. albiflora Schneid. L. bicolor Turcz. &Upsilon. Sieboldii (Miq.) Maxim. in Acta Hort. Petrop. 2: 356 (1873), pro parte-Matsumura in Bot. Mag. Tokyo 16: 70 (1902), excl. f. sericea Matsumura; Ind. Pl. Jap. 2: 267 (1912), pro parte. L. racemosa Dippel, Handb. Laubholz. 3: 720 (1893). L. penduliflora (Oudem.) Nakai in Bot. Mag. Tokyo 37: 79 (1923), excl. quoad specim. cit. Nakai 5578, Nakai 9823, Imai-Ohwi, Fl, Jap., 679 (1957), excl. var. sericea (Matsum.) Ohwi, var. albiflora (Sieb. ex Miq.) Ohwi-Kitamura & Murata, Col. Ill. Herb. Pl. Jap. 2: 99 (1961), excl. var. sericea (Matsum.) Ohwi et L. japonica L. H. Bailey in syn. Japanese name. Miyaginohagi (Hort.; Kaibara, 1698; Matsumura, 1902). A perennial plant, 1-1.5 m high in late summer and early autumn. Stem dependent later, 0.3-0.5 m high in the first year, rarely branched, terete, slightly angular with sparsely ascending appressed hairs (hairs 0.3-0.6 mm long, whitish); aerial part (except the basal part ca. 5 cm from ground level) annual; basal part with axillary and adventitious winter buds. In the second year a few branches emerge from the basal part of the stem, 1-1.5(-2) m long, similar to the stem of the first year, but branched or rarely branched in upper part; most of the aerial part annual; the basal parts (near ground level) with axillary and adventitious winter buds. Branches in subsequent years (5-20 or more in number) coming from the basal parts of stem and branches every year and the number of branches increases according to the size of the stock. Leaves trifoliolate, petiolate, stipulate, spirally arranged. Stipules free, linear- triangular to linear, 2-7 mm long, brownish, persistent. Petioles 1-6 cm long, hairy like the young branch. Rachides 5-30 mm long, similar to the petiole. Terminal leaflets (at middle parts of branches) petiolulate; petiolules 1-3 mm long, swollen; lamina 3.5-9 cm long, 2-5 cm wide, entire, narrowly elliptic, cuneate at the base, acute at the apex (the apex itself with a point), upper surface glabrous (except along the midrib) at anthesis, lower surface appressed-sericeous (hairs 0.5 mm long); lateral ones similar to the terminal ones but somewhat smaller. Inflorescence axillary racemous, one per axil, 10-15 cm long including the peduncle, somewhat compactly to loosely 10-14-flowered; peduncles 1-2(-4) cm long, hairy like the young branch. Primary bracts ovate, about 1 mm long, pubescent, persistent; secondary bracts similar to the first ones. Flowers 12.5-15.4mm long at anthesis; pedicels 2-5 mm long, terete, curved, tomentose. Bracteoles at the base of the calyx narrowly elliptic to ovate, 1-1.5 mm long, pubescent outside, glabrous inside, persistent. Calyx 5.3-5.4 mm long, campanulate four-lobed above the middle part, densely appressed pubescent; tube 1.8- 1.9 mm long; the lower lobe longest; lateral ones 3.4-3.6 mm long, 1.1-1.2 mm wide, triangular-lanceolate, acute at the apex but not acuminate; upper one can be slightly two-clefted. Standard shorter than keel-petal and longer than wings, lamina narrowly elliptic, round at the apex with or without a point, slightly inflexed-auriculate near the base, reflexed in anthesis from near the base. Wings deeper red-purple than standard, lamina narrowly obovate, auriculate at upper basal part, tapering at lower basal part. Keel-petal paler than wings, lamina narrowly obovate. Stamens 10, nearly the same length, diadelphous. Anthers elliptic, shallowly retuse at the apex, ca. 0.7 mm long; before anthesis yellow, sometimes abortive. Ovary elliptic, pubescent, stalked; style pubescent at the basal part, glabrescent at the apical part. Fruits rarely compressed, elliptic, slightly to densely pubescent or nearly glabrous, stalked. Seeds reniform. Cultivated widely as an ornamental plant. Representative specimens. CULTIVATED IN JAPAN. (Siebold s.n., L 908.118-2391-Lectotype of L. Sieboldii Miq.; Thunberg? s.n., L 908.118-2353); Culta in H. B. (Siebold s.n., L 908.118-2356-Syntype of L. Sieboldii Miq.); Nagasaki (Oldham 329, L 908.118-2383); (Buerger s.n., L 908.118-2392 -Syntype of L. Sieboldii Miq.). AOMORI. Naigyushi (cited as "Nadeshiko" by Nakai, 1927) (Kinashi s.n., TI). Cultivated in Botanical Gardens, Koishikawa, University of Tokyo, Tokyo (Akiyama 170, TI). Usually this species is treated as conspecific with L. patens. It is, however, noticeable that the former is different from the latter; L. Thunbergii is very vigorous in the region where L. patens is not distributed; on the other hand, cultivated L. patens does not grow well. L. Thunbergii bears only a few seeds, branches are much dependent, the color of its leaves is dark green, the wing is not as short as that of L. patens. It should be noted that the origin of L. Thunbergii might be a hybrid between L. patens and other species. The flowering season of L. Thunbergii is earlier than that of other species except for L. patens, L. Buergeri, and L. Maximowiczii. As L. Buergeri has whitish yellow standard and keel-petals, a hybrid between L. patens and L. Buergeri is expected to have paler red-purple flowers like those of L. × cyrto-Buergeri and L. × kagoshimensis. But the flowers of L. Thunbergii are not paler red-purple. The shape and color of leaves of L. Thunbergii are similar to those of L. Maximowiczii. The possibility remains that L. Thunbergii has a hybrid origin between L. patens and L. Maximowiczii. However, this possibility is difficult to prove because there is no effective way to detect it at present. For example, the somatic chromosome number of L. patens, L. Thunbergii, and other species of sect. Macrolespedeza is 22 so far as has been examined. We attempted to create artificial hybrids but failed, because when the flowers were covered with fine nets to avoid pollinators, they lost vigor and fell down. |