CHAPTER 2
Shovelling and its Components
|
After he ( Apart from the reasonability of As yet there seem to be few studies which quantitatively deal with the correlations between shovelling and its component characters on the lingual surfaces of the incisors and/or canines. But the observational investigations by Sakai et al. (1965, 1967) on the relations between them on the dentinoenamel junction and by Suzuki and Sakai (1966) on the enamel surface are noteworthy. Although the present author (Mizoguchi, 1978) previously analyzed the relations between these characters by the method of path coefficients, the overall relations between them will be investigated, further including metrical characters such as the depth of the lingual fossa in the present study, by a method different from the previous one, i.e., the principal component analysis method. In this work, two different samples will be analyzed separately by the same procedure to assess the reliability of the results obtained.
|

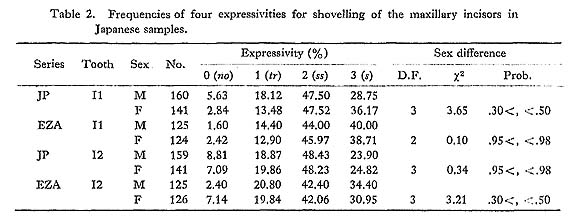
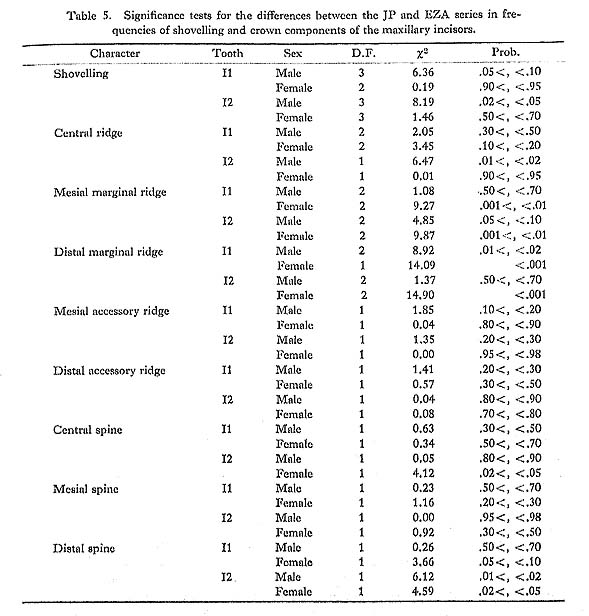

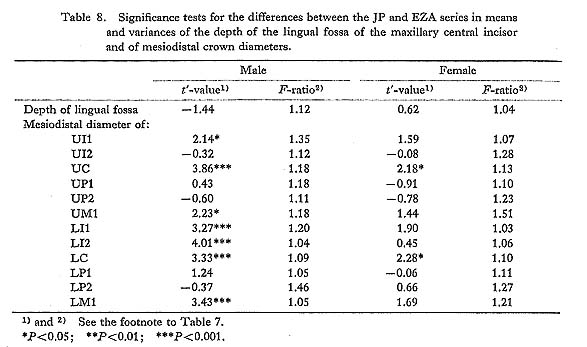

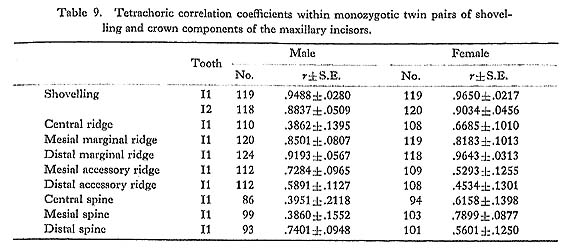
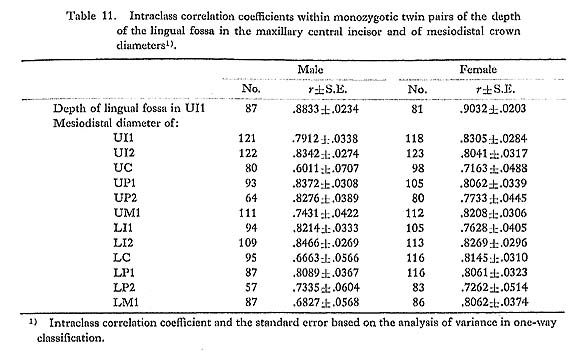


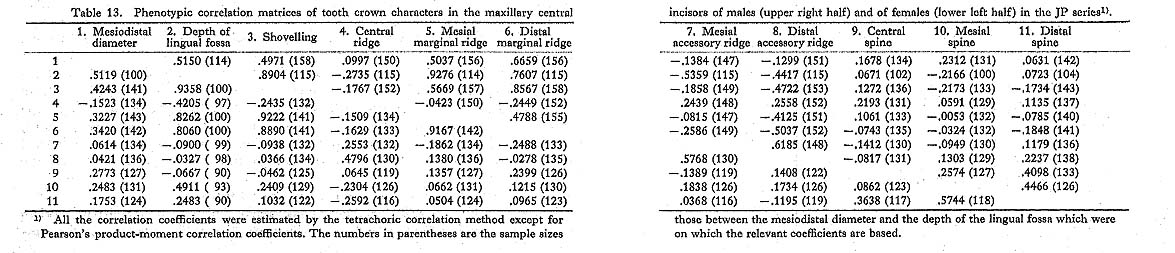 Table 13. Phenotypic correlation matrices of tooth crown characters in the maxillary central incisors of males (upper right half) and of females (lower left half) in the JP series1). |
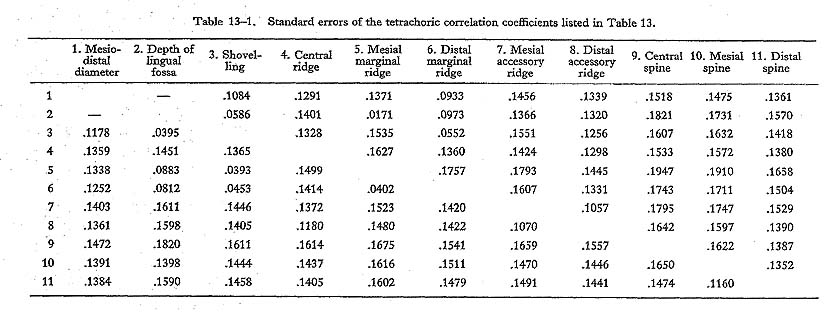

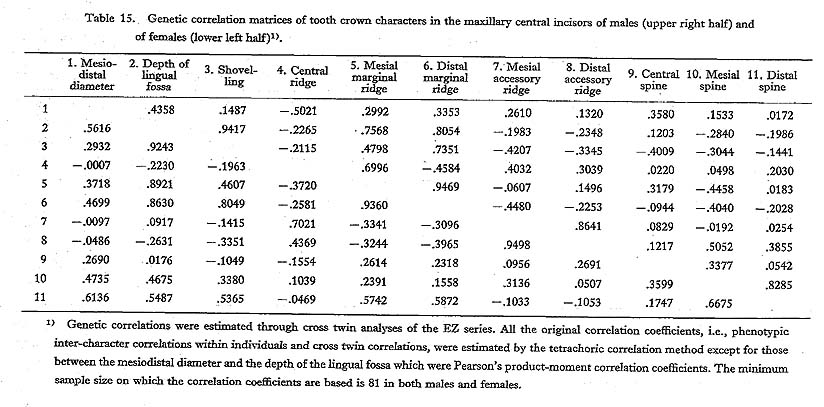
The findings throughout the four phenotypic and two genetic correlation matrices are as follows. First, shovelling and the depth of the lingual fossa had high correlation coefficient estimates which were more than about 0.9, as expected. Secondly, the correlation coefficients of shovelling to the mesial and distal marginal ridges were, again as expected, positive and relatively high, with the latter tending to be higher than the former. Thirdly, the correlation between shovelling and the central ridge was negative, though low. And, finally, shovelling was not so strongly correlated with the mesiodistal diameter, as all the estimates of the correlation coefficient were less than 0.5.
2.3.3. Factor Structure within the Maxillary Central Incisor
The results of the principal component analyses on the above six correlation matrices will be described here. In Tables 16 to 21, the correlation coefficients between the principal components obtained and the original variables, i.e., the characters within the maxillary central incisor, are shown. In factor analysis terminology, the matrix which is composed of such correlation coefficients is called a "factor structure matrix" or "factor loading matrix." As was stated previously, only the principal components whose cumulative proportion reached more than 80% were considered in the present study. The number of principal components obtained in such a way is five in the cases of the phenotypic correlation matrices of JP males as well as of EZ males and females, and four in the case of the phenotypic correlation matrix of JP females, as well as in the cases of the genetic correlation matrices of both sexes. Among these principal components, only the first was strongly correlated with shovelling in every case. In a further search for the principal components to which the contribution of a character is more than about 10%, i.e., those components which have factor loadings of over 0.3 in absolute value, another important principal component can be found to have a similar pattern throughout most of the six principal component analyses. That is, it is the second principal component, which has positive correlations with the mesiodistal diameter, the central ridge and the spines of lingual tubercle, though such an interrelationship is inconspicuous in the JP females.
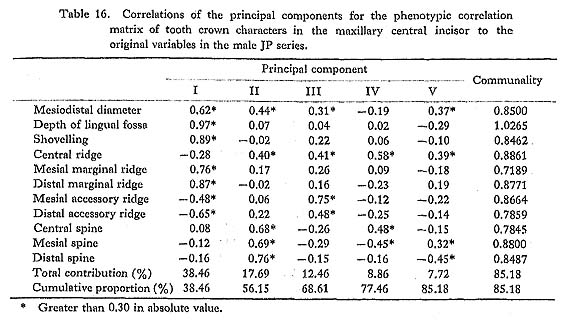
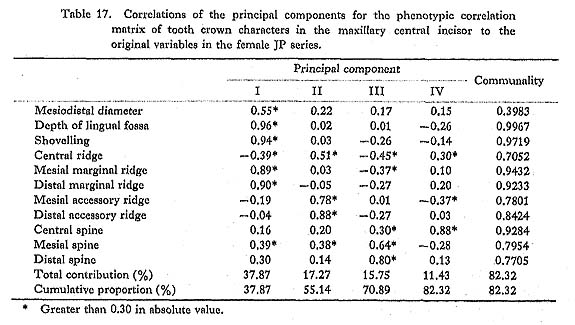

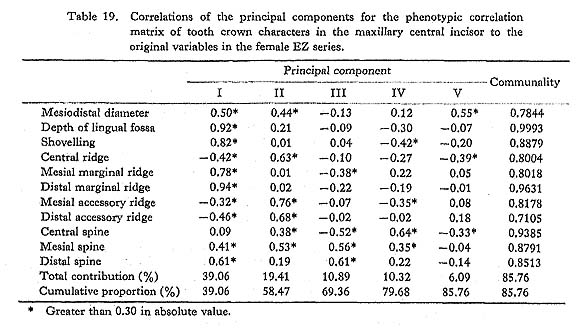
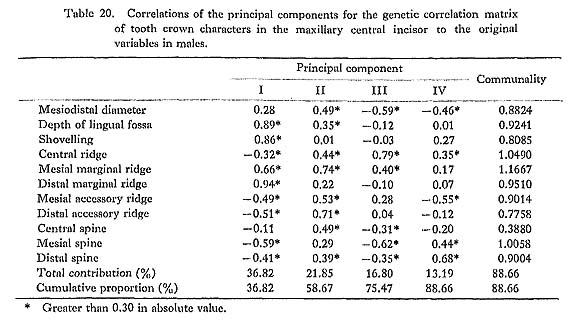
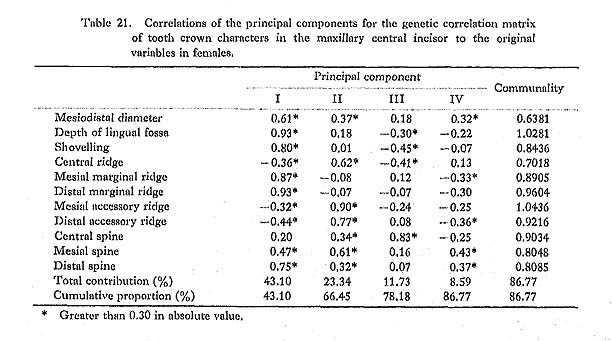
In the present study, however, one of the purposes is to know how each component character of the lingual surface contributes to shovelling in the whole system of interrelations. Therefore, the first principal component will be inspected in greater detail. In Figs. 1 and 2, the degree of contribution, with the factor loading sign, of the first principal component to each character in the JP and EZ phenotypic variations is shown. Similarly, such contributions in genetic variations are illustrated in Fig. 3. From these figures, it is clear that the results of the analyses for the JP and EZ series are nearly the same in spite of their differing in the basic statistics mentioned above. Namely, the first principal component highly contributes to the shovelling and to the depth of the lingual fossa as well as to both mesial and distal marginal ridges in all the cases of the JP and EZ males and females. However, the correlation of this factor with the distal marginal ridge seems to be a little stronger than with the mesial one. The first principal component also has a considerably high correlation with the mesiodistal crown diameter. Further, this principal component has negative correlations, though not strong, with the central ridge as well as with the mesial and distal accessory ridges in every instance.
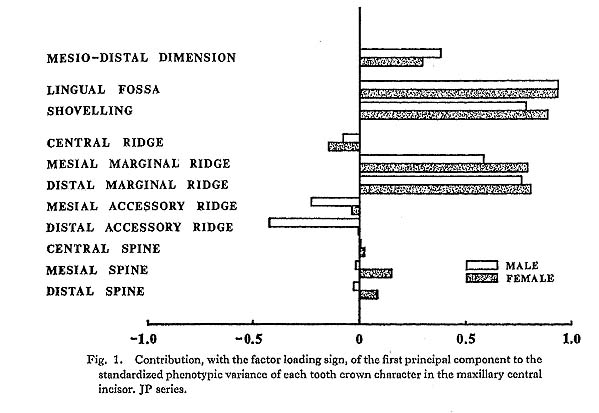
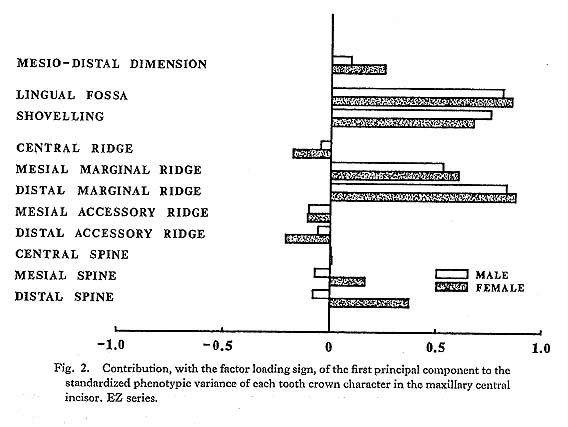

What should particularly be noted in the present study is the interrelationship between the first principal component and the spines of the lingual tubercle. In both the JP and EZ series, the correlation coefficient of the first principal component with the mesial or distal spine in males has the opposite sign to that in females, though the extent of each contribution is not so strong. This trend is much clearer in the first principal component extracted from the genetic correlations (Fig. 3). In other words, it is suggested that there is a qualitative sex difference in the relation of the mesial or distal spine to shovelling, i.e., to the mesial and distal marginal ridges.
The above are the direct results of the principal component analyses. Although these solutions were rotated by the normal varimax method, the resultant solutions were not different in substance before such rotations. Therefore, they will not be described here.
2.4. Discussion
Before the interrelations between shovelling and the component characters of the lingual surface of the incisor are discussed, some problems on the distribution of the depth of the lingual fossa will be considered.
2.4.1. Distribution of the Depth of the Lingual Fossa
It was found that the distribution of the depth of the lingual fossa of the maxillary central incisor tended to have a pronounced tail to the right in one of the four samples (the JP and EZ males and females), if the 5% level was taken as a significance level, or, in two of them at the 10% level (Table 6).
There are several previous reports on the distribution of this character. Hanihara et al. (1970b) examined the distribution of the depth of the lingual fossa of the central incisor in a Japanese sample by the normal-probability paper method and stated that it was almost a completely normal distribution. Mizoguchi (1977b) also suggested that the distribution of this character did not deviate so much from normality because there were no significant differences between the heritability estimates based on the product-moment correlation coefficients and on the tetrachoric correlation coefficients, which could be obtained only under the assumption of complete normality, in all three samples from the Japanese population. Before these, Rothhammer et al. (1968) measured this depth in Chilean populations, which were mixed populations chiefly of Spanish and South American Indian origin, and reported that the significance tests for the deviation from normality showed no significant differences in the central incisor (χ62=12.2, P< 0.10) or in the lateral incisor (χ52=6.5, P<0.30). However, Blanco and Chakraborty (1976), also on the basis of Chilean samples, found the skewed distribution in the central incisor to have a tail to the right (χ52=24.8, P<0.0005). Further, Aas and Risnes (1979) reported that all the central and lateral incisors of the upper and lower jaws had distributions with positive skewness at the 5% level in two Norwegian samples. They also stated that in one of these samples the incisors with the greater means of depth tended to have smaller skewness.
From the above reports and the findings in the present study, it is clear that the distribution of the depth of the lingual fossa has a tendency to tail to the right in a population with a low incidence of shovelling such as Europeans, while it approaches a normal distribution in a population with a high incidence, such as Japanese. This fact strongly suggests that the depth of the lingual fossa is an underlying continuous variate with a certain threshold, i.e., a quasi-continuous variate, and that the threshold is different from population to population.
2.4.2. Contributions of Ridges and Spines
The principal component analyses of the tooth crown characters of the maxillary central incisor based on the two samples, which had different means and different incidences for several characters, showed nearly the same results (Figs. 1-3). Most of these results support the suggestions by some previous authors and increase their validity.
The first principal component was found to have a high positive correlation with shovelling as well as with the depth of the lingual fossa. However, the latter tended to be always slightly higher than the former. This suggests that the depth of the lingual fossa and shovelling are not completely the same character, supporting the view by Dahlberg et al. (1956). The correlation coefficients observed here between shovelling and the depth of the lingual fossa range from 0.89 to 0.94. These are not so different from the correlation coefficient of 0.80 obtained by Turner and Hanihara (1977) on the basis of an Ainu sample. However, the correlation coefficient estimated by Turner and Hanihara appears to be a product-moment correlation coefficient formally calculated between shovelling as a metrical character, of which the expressivities were classified into six grades, and the depth as a continuous variate.
The first principal component which was correlated with shovelling and the depth of the lingual fossa also contributed to both the mesial and distal marginal ridges to an almost equal degree (Figs. 1-3). Further, this principal component always had negative correlations, though not so high, with the central ridge as well as with the mesial and distal accessory ridges. These correlations support the observation by Sakai et al. (1965) and the results of path analyses done by Mizoguchi (1978).
The first principal component also had considerable correlation with the mesiodistal diameter, though the degree of correlation more or less varied from sample to sample. The correlation coefficients estimated are 0.15 to 0.50 between shovelling and the mesiodistal diameter, and 0.36 to 0.52 between the depth of the lingual fossa and the mesio distal diameter. These may be directly compared with the results obtained by Kirveskari and Alvesalo (1979) on the basis of data from Skolt Lapp and Finn populations. They combined samples from the two populations to estimate correlations between the mesiodistal diameters and the depth of the lingual fossa. The correlation coefficient of 0.430 for the maxillary central incisor and that of 0.432 for the lateral one are both within the range observed in the present study. Mizoguchi (1978) reported correlation coefficients of 0.20 to 0.36 between shovelling and mesiodistal diameter in the maxillary central incisor and of 0.35 to 0.41 in the lateral incisor, which were estimated by the tetrachoric correlation method on the basis of samples which partly overlapped with the present samples. These correlation coefficients are also within the range observed here. After all, the correlation between shovelling or depth of the lingual fossa and the mesiodistal diameter does not seem to be so strong (being as high as about 0.4). Dahlberg et al. (1956) had already suggested such a relationship and Suzuki and Sakai (1966) had confirmed it by using qualitative statistical methods.
A new finding in the present study is that for males and females the way in which the mesial and distal spines of the lingual tubercle contribute to shovelling is different. This was ascertained in two independent samples (Figs. 1 and 2) and most definitely indicated in the results of the principal component analyses of genetic correlations (Fig. 3). That is, the correlation of the first principal component, which is highly correlated with shovelling, to the mesial or distal spine has a tendency to be negative in males but positive in females. Although the high contributions of the mesial and distal spines to shovelling in the female lateral incisor have already been pointed out by Mizoguchi (1978), such a qualitative between-sex difference as shown in this study had been previously unknown.
In order to interpret this relationship of the spines to shovelling, previous literature on shovelling and the lingual tubercle will be presented in the next section.
2.4.3. Shovelling in Fossil Man
The history of investigations on the relations between shovelling and the lingual tubercle is intertwined with the history of the dental morphology of fossil man, such as Pelting man, Neanderthals, etc., by Weidenreich (1937), Adloff (1938) and others. Of course, some workers have also statistically examined the relations between them on the basis of data from modern man.
Suzuki and Sakai (1966) confirmed, using the χ2-test, that tnere was a significant parallelism in the development of shovelling and the lingual tubercle of the maxillary central incisor of modern Japanese (P<0.01). They also stated that the spines of the incisors were less developed than those of the canines and seldom made the lingual fossa narrow or full, but rather often made the boundary between the lingual fossa and the basal part of the tooth crown clearer. However, it is unknown whether the data used for this statement were from males or females. Moreover, their results are not comparable with those of the present study because they did not distinguish the sides on which the spines occurred, i.e., mesial, distal or central. But their results at least seem to be evidence to prove that shovelling or the lingual tubercle relies on the existence of the other.
Hanihara (1970b) investigated the interrelations between the morphological variation of the maxillary lateral incisor and the mesiodistal crown diameters of all the permanent teeth except the third molars on the basis of a Pima Indian sample. He initially classified the lateral incisors into three types: reduced or conical type; malformed or over-developed type, including barrel-shaped teeth with a highly developed lingual tubercle; and normal type. However, transitional types such as peg-shaped teeth were excluded from his analysis. As a result, he found that in male individuals who had reduced lateral incisors, the other teeth (UI1, LI2, LM2) were also smaller at the 5% significance level than those of individuals with normal lateral incisors, while in individuals with lateral incisors of the over-developed type, the other teeth (UI1, UP1, UM1, LM1) were significantly larger. In the latter case, however, the size of the maxillary lateral incisor itself did not significantly differ from that of the normal type group. In females, no teeth but the maxillary lateral incisor of the reduced type group were significantly different from those of the normal type group, while on the other hand all teeth including the maxillary lateral incisor of the over-developed type group were significantly larger than those of the normal type group. Hanihara (1970b) inferred that the occurrence of these reduced or over-developed lateral incisors was associated with the determination of the overall size of dentition. If most of the incisors of the over-developed type have well-developed lingual tubercles, his findings may also be interpreted to show that the lingual tubercle is closely related to the mesiodistal crown diameter. The second principal component extracted from the phenotypic correlation matrices in the present study was correlated with both the mesiodistal crown diameter and spines of the lingual tubercle in three of the four analyses (Tables 16-19). If only the factor loadings of more than. 0.4 in absolute value are taken into account, this component is considered to be intensively related to the mesiodistal diameter, the central ridge and the three spines in JP males; to the mesiodistal diameter, the mesial and distal accessory ridges and the three spines in EZ males; and to the mesiodistal diameter, the central ridge, the mesial and distal accessory ridges and the mesial spine in EZ females. This suggests that there is a considerable amount of variation which is caused by a common factor in the lingual tubercle and the mesiodistal diameter but which is independent of the shovelling. This is compatible with Hanihara's (1970b) finding stated above.
Additionally, there are some other investigations on the associations between the lingual tubercle and shovelling based only on morphological observations.  (1921) stated that the lingual tubercle might occur in both shovel-shaped incisors and incisors with a flat lingual surface in modern man. Moorrees (1957) claimed that there was no association between shovelling and the lingual tubercle because Aleut maxillary incisors which were shovel-shaped had no lingual tubercles. On the other hand, Dahlberg et al. (1956) said that better-developed lingual tubercles tended to occur in shovel-shaped incisors. As already mentioned above, however, a certain relationship between these two characters has been confirmed.
(1921) stated that the lingual tubercle might occur in both shovel-shaped incisors and incisors with a flat lingual surface in modern man. Moorrees (1957) claimed that there was no association between shovelling and the lingual tubercle because Aleut maxillary incisors which were shovel-shaped had no lingual tubercles. On the other hand, Dahlberg et al. (1956) said that better-developed lingual tubercles tended to occur in shovel-shaped incisors. As already mentioned above, however, a certain relationship between these two characters has been confirmed.
In passing, Dahlberg (1951) stated that barrel-shaped teeth with exceedingly devel-oped lingual tubercles did not occur in individuals who did not exhibit shovelling, and, therefore, the barrel shape was associated with shovelling. Later on, some workers regarded this barrel shape as extremely developed shovelling. For example, Baume and Crawford (1980) classified the expressivity of shovelling into five grades: absence, moderate trace, marked, double and barrel. Against such a view, Mayhall (1979) insisted, on the basis of Canadian Eskimo dentition, that there was no continuity between shovelling and the barrel shape. These arguments are dependent upon the definitions of both shovelling and barrel shape. Since the present author's view was stated in the previous section (see 1.2.), it is not repeated here.
Adloff (1938) distinguished two different types of incisors, "Schaufelform" and "Höckerform," according to the development of the lingual tubercle, and claimed that the maxillary central incisors of such fossil men as Peking man were not "Schaufelform," which occurs only in modern man. However, Weidenreich (1937) and Robinson (1956) did not accept his definition of shovelling, which involved the presence or absence of the lingual tubercle. The present author holds the same opinion as Weidenreich and Robinson, but, apart from the definition, does not think that Adloff's view was necessarily wrong. It seems right, at least after the evolutionary stage of Homo erectus, that the shovel-shaped incisors seen in modern man were derived secondarily from incisors with well-developed lingual tubercles through the reduction of those lingual tubercles. In the following paragraphs, evidence for the interrelationship between the lingual tubercle and shovelling will be presented in the dentition of modern and fossil man.
First, in the case of living modern man, both the maxillary central and lateral incisors generally have marginal ridges which converge towards the cervix of the tooth, regard-ess of their development, and have a triangular outline of the lingual surface even if the ingual tubercle is highly developed or cusp-like (Pl. I, , 1920; Plate 13, Dahl-berg, 1951; Fig. 12, Moorrees, 1957). Besides, the shape of the lingual cusp of modern nan, even of barrel-shaped incisors, seems considerably different from those in the ncisors of Krapina man or the like (Plate 13, Dahlberg, 1951; Fig. 6B, Dahlberg, 1963a; Fig. 1, Hanihara, 1970b).
An outline of the previous reports on shovelling and the lingual tubercles of historic and prehistoric Homo sapiens sapiens, including work only on the incidence of shovelling, will be presented below.
An investigation of over 300 Indian skulls from Pecos Pueblo (about A.D. 1100) howed that ss and more developed shovelling occurred at a rate of 89.5% in the maxil-ary central incisors, in which only two individuals had incisors with a triangular well-leveloped lingual cusp (Nelson, 1938). These incisors were shovel-shaped, but the ingual cuspa reached to the three-quarter point of the crown and were fused with the ingual surface. These lingual tubercles seem considerably different in shape from hose of Homo erectus and Homo sapiens neanderthalensis. In Chilean Diaquitas Indians of about the 10th century, the incisors with lingual fossa over 0.3 mm deep were found o make up 80.3% of the population (Campusano et al., 1972). All of the incisors exca-ated from the three prehistoric Ohio Valley sites, i.e., Glacial Kame of about 2000-500 B.C., Adena of about 1500 B.C.-A.D. 300 and Ohio Hopewell of about 100 B.C.-A.D.400 showed tr or more developed expressivities of shovelling in both the maxillary entral and lateral incisors (Sciulli, 1979).
About 93% of the maxillary incisors excavated from the Hane Dune site, Marquesas, f A.D. 1110-1635 and 82% of the maxillary incisors from the Nebira site, Papua New Guinea, of A.D. 1000-1600 exhibited tr or more developed shovelling (Pietrusewsky, 976).
In the maxillary central incisors from Bali, Indonesia of the Bronze Age, the frequency of shovelling of ss and more developed expressivity was found to be 38.9%, and, in the lateral incisors, to be 26.2% (Jacob, 1967). Jacob recognized lingual tubercles in four of approximately 80 maxillary incisors. The teeth with lingual tubercles were not shovel-shaped.
In An-yang Chinese who lived 3100 years ago, the frequency of s shovelling was found to be 43.6% (Turner, 1976). In the Jomon people of Japan, however, no shovel-shaped maxillary central incisor with s expressivity was found (Turner, 1976). As regards the Jomon incisors, Turner (1979) recorded the frequency of shovelling in more detail, where ss and s were 29.6% or tr and more developed expressivities 100% for the maxillary central incisor, and ss and s were 34.6% or tr and more developed expressivities 100% for the maxillary lateral incisor, Gates (1956) said that nearly all the Japanese skulls studied from the Kamakura period, about 700 years ago, had shovel-shaped incisors. According to his Tables 4 and 5, the frequency of shovelling, maybe of over tr, is 83.3% (ten out of twelve skulls) in males and again 83.3% (five out of six skulls) in females. He also observed shovelling in at least two of the four Jomon skulls excavated at Sanganchi near Sendai, Japan. The preliminary calculations based on Mizo-guchi's (1982) data and the unpublished data of the present author showed that the incidence of ss plus s shovelling was 88.9% (eight out of nine individuals) in UI1 and 90.0% (nine out of ten) in UI2 of the Kofun period, from about the third to 8th/9th century A.D., excavated from the Kanto district of Japan. Taking account of tr and more developed expressivities, all the Kofun incisors are characterized by shovelling. Another preliminary calculation based on Hiramoto and Mizoguchi's (n.d.) data and the unpublished data of the present author on Jomon incisors excavated from the Chubu, Kanto and Tohoku districts of Japan showed that the frequency of ss plus s was 91.7% (11 out of 12 individuals) in UI1 and 93.3% (14 out of 15) in UI2. In this case also, the total incidence of tr, ss and s was 100% in either UI1 or UI2.
Weidenreich (1937) stated that the maxillary central incisors of Upper Cave man from Choukoutien, China, resembled those of Peking man. Observing fig. 9 in Weidenreich (1937), however, the cervix of the former seems narrower and, besides, the lingual tubercle seems to be slightly less developed.
In a young adult male skeleton excavated near Chéria, Algeria, dated to 9130 B.P., both the right and left maxillary lateral incisors exhibited tr shovelling (Meiklejohn et al., 1979), From the Mesolithic site Wadi Halfa, in Sudan, which dates back 8000-11000 years ago, at least nine incisors were excavated (Greene et al., 1967), of which two (22.2%) were shovel-shaped incisors of ss or more developed expressivity, or, taking tr and more developed expressivities into account, seven (77.8%) were shovel-shaped. In most of the maxillary incisors the lingual tubercle was present. From Fig. 3 of Greene et al., it seems that the cervices of these teeth are narrow or are those of modern man.
In the Merovingian site, about the 5th to 8th century, at Vexin in the northern part of France, ss shovel-shaped incisors were found to constitute 4.76% of the maxillary central incisors, and tr to be 1.48% of the maxillary lateral incisors (Ménard, 1978). From the Bronze Age sites at Fleuré (Vienne) and Puyraveau (Deux-Sèvres) in the western part of France, 244 maxillary central incisors and 197 maxillary lateral incisors of the right and left sides were excavated. Shovelling of tr or more developed expressivities was recognized in 19% of the centrals and in 29% of the laterals (Patte, 1976). According to Patte (1976), Brabant found shovel-shaped incisors to be 27% of the maxillary central incisors and 25% of the laterals from Manthelan (Indre-et-Loire) dolmen, and 16% of both maxillary central and lateral incisors from Moncouè (Thouarsais) dolmen. Brabant (1971) investigated the teeth from various Megalithic sites in France dating from about 2500 to 1000 B.C. and reported the results together with those of previous work. According to him, shovel-shaped incisors of ss or more developed expressivity were 29.4%, of the maxillary central incisors and 37.8% of the lateral incisors from Neolithic sites in France; 20.6% of the centrals and 47.0% of the laterals from Neolithic sites in Switzerland; 30,8%) of the centrals and 35.1% of the laterals from Megalithic sites in France; and 19.2% of the centrals and 28.3% of the laterals from medieval sites in Belgium. Brabant compared these with the frequencies in modern whites and claimed that in Western Europe the incidence of shovelling had decreased since the Neolithic Age.
In the 26 maxillary central incisors from a site at Knossos, Crete, which is of the Middle Minoan III period dating back to about 1750-1550 B.C., the frequency of shovelling of ss or more developed expressivity was found to be 73%, or, taking tr or more developed expressivities into account, 92%, (Carr, 1960). Carr calculated Penrose's size and shape distances between this site and other selected populations based on the mesio-distal diameters of tooth crowns. The Knossos teeth were smaller than those of not only Australian Aborigines but also Pecos Indians, Eskimos, Javanese, Bantu, American whites, etc. The size distances showed these teeth to be most similar to those of Lapps and Bushmen, but the shape distances showed that they were nearest to American whites and Pecos Indians. Carr (1960) stated in conclusion that the Knossos teeth were, as a whole, most similar to those of American whites and of Lapps, as had been expected.
The above is a description restricted only to Homo sapiens sapiens. The figures indicating the frequencies of shovelling reported by previous authors cannot be compared directly with one another because the criteria of observation adopted by those investigators may be considerably diverse. However, the following trends may be discerned.
The incidence of shovelling was high in the northern part of China about 3000 years ago, but not so high in Japan at almost the same time. It was not so high in Indonesia about 2000 years ago either. In the New World a high incidence of shovelling has continued for the past 3000-4000 years. In Europe, the incidence was considerably higher 3000-4000 years ago, but has since decreased. In Africa, it has been consistently low for the past 10,000 years or thereabouts. As regards the lingual tubercle, there is no evidence to show that this character seen in the fossil remains of Homo sapiens sapiens was particularly different in shape or in incidence from that in living modern man.
Next, earlier reports on the teeth of Homo sapiens neanderthalensis will be examined. First,  -Kramberger (1906) pointed out that Krapina man from Yugoslavia had maxillary incisors with well-developed marginal ridges and a deep lingual fossa as well as a thickened tubercle at the basal part of the crown. He also observed that this lingual tubercle was sometimes divided into two or three portions by small grooves and that in the case of the lateral incisors the tips of the lingual tubercle might be separated from the lingual surface. According to the photographs (Figs. 34 and 35) in -Kramberger (1906), the Krapina incisors were different from those of modern man. In both central and lateral incisors of Krapina man, the mesiodistal width at the cervix of
the tooth crown is almost the same size as the mesiodistal width near the incisal edge, and both marginal ridges run almost parallel to one another. These features seem to be related to the large width of the lingual tubercle itself, as was pointed out by Robinson (1956). Recently, Wolpoff (1979) described the Krapina teeth in more detail. The abundant photographs included in his report clearly show the above-mentioned points.
-Kramberger (1906) pointed out that Krapina man from Yugoslavia had maxillary incisors with well-developed marginal ridges and a deep lingual fossa as well as a thickened tubercle at the basal part of the crown. He also observed that this lingual tubercle was sometimes divided into two or three portions by small grooves and that in the case of the lateral incisors the tips of the lingual tubercle might be separated from the lingual surface. According to the photographs (Figs. 34 and 35) in -Kramberger (1906), the Krapina incisors were different from those of modern man. In both central and lateral incisors of Krapina man, the mesiodistal width at the cervix of
the tooth crown is almost the same size as the mesiodistal width near the incisal edge, and both marginal ridges run almost parallel to one another. These features seem to be related to the large width of the lingual tubercle itself, as was pointed out by Robinson (1956). Recently, Wolpoff (1979) described the Krapina teeth in more detail. The abundant photographs included in his report clearly show the above-mentioned points.
The teeth associated with the early Upper Paleolithic or Aurignacian culture from Vindija cave, Croatia, Yugoslavia, were the subject of a recent report by Wolpoff et al. (1981). Among these teeth there is one maxillary central and one lateral incisor, each of which has highly-developed shovelling and a strong lingual tubercle. They closely resemble the incisors of Krapina man.
The two maxillary incisors which were found near the child mandible of Ehringsdorf in Germany also have well-developed shovelling and lingual tubercles in them and have been said to closely resemble those of Krapina man (, 1930; Weidenreich, 1937;Patte, 1962).
The maxillary incisors of La Quina (, 1930; Patte, 1962) and Le Moustier (Weidenreich, 1937; Patte, 1962) in France again have highly-developed shovelling and lingual tubercles. The skeletal remains of Chaâteauneuf 2 associated with the Mousterian culture in France also have maxillary central and lateral incisors with very strong shovelling and large lingual tubercles (Tillier, 1979). According to Tillier's photograph (Planche II), both marginal ridges of the central incisor slightly converge towards the cervix. This condition seems to be halfway between that of modern man and Krapina man.
McCown and Keith (1939) described the teeth of Tabùn man and Skhùl man associated with the Levalloiso-Mousterian culture from Mount Carmel, Israel and suggested that the former had more Neanderthal and the latter more modern characteristics. The maxillary central incisor of Tabùn man (Tabun Serie III, left UI1) has a well-developed lingual tubercle whose tip tends to be separated from the lingual surface, resembling the incisor of Le Moustier more than that of Krapina man in this point (McCown and Keith, 1939). In a figure from McCown and Keith (Fig. 121, C), the shovelling seems to be ss, and both marginal ridges seem to have a tendency to slightly converge towards the cervix. The two maxillary lateral incisors of Tabùn man also have highly-developed shovelling and cusp-like lingual tubercles, and are Neanderthal-like (McCown and Keith, 1939). Judging from those figures (Fig. 122, A and C), both marginal ridges seem to converge towards the cervix in one of these lateral incisors, as in the case of modern man, but, in the other lateral incisor, to run almost parallel to oneanother. McCown and Keith stated that the Tabùn lateral incisors also resembled those of Le Moustier. On the other hand, the unerupted maxillary central and lateral incisors of Skhùl man, whose dentition has been said to be like that of modern man, show well-developed shovelling and lingual tubercles (McCown and Keith, 1939). Judging from that figure (Fig. 137, B), both marginal ridges of the central incisor run nearly parallel to each other, and, therefore, seem to the present author to be unlike those of modern man.
There are skeletal remains from Qafzeh (Homo VI), Israel, associated with the Mousterian, which have been said to be morphologically in the middle of Classic Neanderthals in Western Europe and so-called Homo sapiens, modern man (Vallois and Vandermeersch, 1972). In reference to this case, Vallois and Vandermeersch stated that the skull of this young male had large incisors with no shovelling. According to their photographs (Figs. 10 and 12), however, shovelling (tr or ss) seems recognizable in the maxillary central incisors, but certainly not so highly developed as in those of Krapina man. His lingual tubercles seem not to be so strongly developed either.
Amud I, excavated from Israel, has been thought to be a Neanderthal who lived in a transitional period from the Middle Paleolithic to the Upper Paleolithic (Suzuki and Takai, 1970). The size of the maxillary central incisors of Amud I was said to be in the range of those of Neanderthals. The shovelling was inferred to be less developed, though this was not so certain because of dental attrition (Sakura, 1970). The lingual tubercles in the maxillary central and lateral incisors were reported to be well developed. According to a photograph (Plate 35), however, the lingual tubercles, especially of the central incisors, seem not so well developed as those of Krapina man.
A Mousterian boy from Teshik-Tash cave in the Central Asian part of the USSR has been considered, based on the cranial morphology, to be from a transitional stage between European Classic Neanderthals and Skh ù l man of Israel (Weidenreich, 1945b). His maxillary lateral incisors were extremely shovelled, so Weidenreich considered it an indication of the trend in Mongoloids.
The maxillary central and lateral incisors of Tingtsun man from China have highly-developed shovelling and lingual tubercles (Woo, 1956; Chang, 1962). What is called Ordos man is known by only one left maxillary lateral incisor, which has definite shovel-ling and a well-developed lingual tubercle (Woo, 1956; Chang, 1962), Observing the teeth of the above Tingtsun and Ordos man in Fig, 2 of Chang (1962), the lateral incisors of both Tingtsun and Ordos man seem to have marginal ridges tending to converge towards the cervices. In this point, they appear like those of modern man. However, the marginal ridges in the central incisor of Tingtsun man seem to run almost parallel to one another.
Rabat man from Morocco, considered to be a contemporary of those who lived during the Riss glacial stage in Europe, has been said to have large dentition as a whole and incisors with strongly developed lingual tubercles and less developed shovelling (Thoma and Vallois, 1977). Thoma and Vallois said that the maxillary first and second molars had well-developed hypocones, and that the premolars were strongly molarized. According to their figure (Fig. 1), the lingual cuaps of the maxillary firat and second premolars certainly tend to be divided into two parts. However, shovelling of the maxillary central incisors seems close to ss rather than to tr, and that of the lateral incisors also seems to be ss. Further, the marginal ridges of the lateral incisors appear to slightly converge towards the cervices.
The above is a reexamination of the morphology of the Neanderthal maxillary incisors on the basis of descriptions by previous authors, and may be summarized as follows.
First, European Neanderthals such as Krapina, Vindija, Ehringsdorf, La Quina, Le Moustier, Ch â teauneuf, etc. generally had maxillary incisors which are characterized by a deep lingual fossa and a highly developed lingual tubercle. The tooth crowns of these teeth are as wide mesiodistally near the cervix as at the incisal edge, and the lingual tubercle flows into the lingual fossa between the two parallel marginal ridges. In the lateral incisors the lingual tubercle sometimes tends to have tips which are separated from the lingual surface. Similarly, Tingtsun and Ordos man from China had incisors with most of the above characteristics, but the necks of the lateral incisors are slightly narrower, i.e., like those of modern man. On the other hand, the Neanderthals from Israel, especially Qafzeh and Amud man, are said to have been morphologically transitional types becoming modern Homo sapiens. The shovelling of the teeth is weakly developed, and the lingual tubercles are not so developed as those of European Neanderthals. Among the Mount Carmel specimens, also from Israel, there are said to be European Neanderthal and modern man types. It is certain that some of their lateral incisors had narrow necks like those of modern man. Rabat man from North Africa, who is considered to be chronologically older than European Neanderthals, had incisors with less developed shovelling and, at the same time, well-developed lingual tubercles, as well as large posterior teeth.
Some interrelationship between the morphology of the incisors and the geographical distribution of human populations seems to be suggested through the above examination. Before going into detail on this point, however, we will further examine the morphology of the incisors of earlier hominid remains.
Peking man was asserted by Weidenreich (1937) to be closer to the Mongoloid than to any other stock mainly because of his maxillary incisors which had highly-developed shovelling. The maxillary central incisors are characterized by a strongly developed lingual tubercle in addition to deep shovelling. The two lateral incisors preserved have well-developed shovelling but no definite lingual tubercles, and therefore the crowns of these teeth are not so different in shape from those of modern man.
Observing the replicas of the two maxillary central incisors of Yuanmou man discovered in South China, which are housed in the National Science Museum, Tokyo, it is seen that they have well-developed shovelling and lingual tubercles as those of Peking man do. Both lingual marginal ridges run parallel to one another.
The right maxillary lateral incisor, found near a skull of Java man, Skull IV, from Sangiran (Weidenreich, 1945a) is also a shovel-shaped incisor (van Heekeren, 1972). According to Plate 5 in Weidenreich (1945a), the shovelling seems to have ss expressivity, but the lingual tubercle appears to be undeveloped. In these points, this tooth appears to resemble the lateral incisors of Peking man.
Summarizing the above, while the central incisors of Homo erectus are extremely similar to those of European Neanderthals, the lateral incisors seem rather more like those of modern man.
Finally, the teeth of Australopithecus are reviewed. Robinson (1956) studied these in detail, as mentioned above. The nine maxillary central incisors of A.robustus excavated from Swartkrans all have moderately-developed shovelling, weakly swollen lingual bases of the crowns and mesial and distal marginal ridges which converge towards the cervices. The two central incisors of A.africanus from Sterkfontein also have moderately-developed shovelling and no so-called lingual tubercles. However, they have wealdy-developed spines from the lingual base of the crown. With respect to the lateral incisors, those of A.robustus from Swartkrans and of A.africanus from Sterkfontein as well as from Makapan were described by Robinson. All of them have weak or moderate shovelling and lack developed lingual tubercles. Robinson (1956) stated that such morphology of the incisors was also frequently observable in modern man.
Tobias (1967) described the maxillary incisors of Zinjanthropus found in Olduvai Gorge. According to him, a trace of shovelling is recognizable in them despite the heavy dental attrition. Further, the lingual base of the crown is slightly swollen and both marginal ridges converge towards the cervix. Tobias pointed out the similarities between these incisors and those of South African australopithecines reported by Robinson (1956).
Leakey and Leakey (1978) described the hominid fossils excavated from Koobi Fora. Judging from their photographs, the left maxillary central incisor of KNM-ER 1590B, which Leakey and Leakey considered to be Homo habilis, seems to have tr or ss shovelling and marginal ridges which converge towards the cervix. The right maxillary lateral incisor of KNM-ER 1813A, which was identified as Australopithecus africanus, has shovelling probably of ss and also marginal ridges which seem to converge towards the cervix.
Recently, Johanson et al. (1982) described the teeth of Australopithecus afarensis from the Pliocene Hadar Formation, Ethiopia. Among them there are over ten maxillary incisors. According to their description and photographs, the central incisors have relatively definite marginal ridges, which slightly converge towards the cervices. Their shovelling seems to be (tr or ss. The mesiodistal width of the marginal ridges themselves tends to be narrow compared with the mesiodistal crown diameter, being like those of apes. The lingual tubercles are relatively well developed, and some of them resemble those of Homo erectus. The maxillary lateral incisors are considerably smaller than the centrals, and have some other ape-like features. The lingual tubercles are in general weakly developed, and the shovelling seems to be tr or ss.
In conclusion, it seems that both South African and East African australopithecines had incisors with weakly or moderately developed shovelling. In this point, they are much more like modern man than Homo erectus or European Neanderthals.
Based on the above examination, the morphology of the maxillary incisors of the hominids from the australopithecines to modern man may be roughly classified into three types. The first type of incisor has weak or moderate shovelling, a slightly swollen basal part of the lingual surface of the tooth crown and lingual marginal ridges converging towards the cervix. This type ia frequent in australopithecines and most modern human populations. The second type has extremely-developed marginal ridges which run parallel to one another, a very deep lingual fossa and a large lingual tubercle, The central incisors of Homo erectus and European Neanderthals are representatives of this type, The third type also has highly-developed shovelling or marginal ridges, but the marginal ridges converge towards the cervix. Further, this type has no developed lingual tubercle. The lingual tubercle, if any, is in the form of small spines like folds at the lingual base of the tooth crown, and therefore different from those of Homo erectus and European Neanderthals. This third type of incisor is often found in modern Eskimos, Pima Indians, etc.
It seems very difficult to believe that these three incisor types, considerably different in structure, satisfy the same functional demands. To the present author, the causes for such differences seem to be found in the interrelationship of geographical environments, climatic conditions, degrees of cultural development in human populations and. so forth.
In Africa, the maxillary incisors have continued to be the first type mentioned above from australopithecines through Neanderthals and the fossil Homo sapiens sapiens who was living about 10,000 years ago, to modern Africans (Carbonell, 1963; Barnes, 1969). On the other hand, in eastern Asia, Homo erectus and Neanderthals had the second type of incisor, but modern Asians have the first type. In Europe, the Neanderthals who lived during the period from the Riss to the Würm glacial stage had the second type of incisor, but modern Europeans have the first type. The Neanderthals in western Asia, who are said to represent the transitional condition before becoming modern man, had a median type of incisor between the first and second types described above, in which shovelling was less developed while the lingual tubercles were relatively well developed. And the third type of incisor is restricted to the modern Eskimos in the Arctic, as well as to the Indians in the Americas. What does this mean?
Weidenreich (1937) considered that well-developed shovelling was a Mongoloid characteristic throughout evolution, and that the difference between the incisors of modern man and of Homo erectus and Neanderthals was in the degree of development of the lingual tubercles. He maintained that the shovelling of the incisors of Le Moustier and Krapina man was less developed and, therefore, rather more like that of modern Europeans than that of modern Mongols. Thus Weidenreich directly linked Peking man with modern Mongols chiefly because shovelling was highly developed in both populations. Refuting this view, Robinson (1956) argued that because shovelling was recognizable not only in australopithecines, Neanderthals and modern Mongols but also in other modern populations, Weidenreich's hypothesis linking Peking man with the modern Mongols was not necessarily correct. Robinson asserted that shovelling was a character common to all the hominids. In fact, it is certain that the maxillary central incisors of Peking man closely resemble those of European Neanderthals. Therefore, even if Peking man is a direct ancestor of the Mongoloid population, highly developed shovelling is not necessarily evidence for Weidenreich's hypothesis. However, another view by Weidenreich (1937) seems reasonable-that the substantial difference between the incisors of modern man and those of Homo erectus and Neanderthals is in the degree of development of the lingual tubercles. Adioff (1938) thought that "Schaufelform" seen in modern man was derived secondarily by reduction of the lingual tubercle from "Höckerform" with the well-developed lingual tubercle seen in Krapina man, for example. This hypothesis seems to correspond to the post-Neanderthal stage in Europe. This is not inconsistent with the view of Brace (1967, 1979) and Sakura (1970) that the amount of dental reduction from the Neanderthal stage to the present is greater in the anterior than in the posterior teeth. It is a fact that the morphology of the anterior teeth has changed through the period from the Neanderthal to modern man. The proposition that developed shovelling is a characteristic of the Mongoloid is applicable only to modern man.
The evidence above is not enough to explain the causes and evolutionary processes which have produced three such types of incisor. Assessing the phenomena, however, a hypothesis may be postulated as follows. In Africa, the first type of incisor has continued since the emergence of the horainid, In Europe, the first type changed to the second type before or at the stage of Neanderthals, and, later, back to the first type. In Asia, as in Europe, the second type once changed into the first type after the Neanderthal stage, but some Asian anterior teeth developed in a different way from before, presumably as a result of direct or indirect adaptation to a severely cold environment. Such incisors are the third type of incisor observable in the modern inhabitants of the Arctic and Americas.
To clarify the reason the second type of incisor appeared, other evidence and examination will be required. This is one of the problems to be solved in the future. The difference between the third and the first types of incisor is a variation which has taken place in identical modern human populations. The causes for this variation will again be discussed in the final chapter (4.).
Through the above examination, the outline of the variation in morphological structure of the human maxillary incisors seems to have been made clear. The key for explaining the findings in the present study-that the first principal component contributes in different ways to shovelling and to the lingual tubercle in males and females-seems to be the state of the lingual tubercles in large tooth crowns such as those of Homo erectus and Neanderthals. That is, male incisors with the greater mesiodistal crown diameters of modern man seem to have lingual tubercles of which the spines tend to flow directly into the lingual fossa as in the cases of Homo erectus and Neanderthals. On the other hand, female incisors with relatively smaller mesiodistal diameters seem to have lingual tubercle spines which fuse to the bases of the marginal ridges and therefore tend rather to promote the degree of development of shovelling superficially. This morphological condition seems to be a cause for the qualitative between-sex difference in the first principal component. The second principal component, which is independent of shovelling, has high positive correlations with both the mesiodistal diameter and the spines of the lingual tubercle. That is to say, the spines tend to be strongly developed when the mesiodistal diameter is large. This finding is consistent with the fact that not only European Neanderthals with highly developed shovelling but also an African Neanderthal, Rabat man, with less developed shovelling equally have wide incisors with well-developed lingual tubercles. Therefore, the wide incisors appear to demand the existence of the lingual tubercles.
In conclusion, shovelling is inferred to function relatively independently of the lingual tubercle.
2.4.4. Structure and Function of the Incisor
In this section studies by previous authors on the structure and function of the incisors will be reviewed, and the problems to be solved will be pointed out.
Shovelling has since (1920) been thought to be a structure for strengthening the incisors, but this has not been conclusively demonstrated (Mayhall, 1977). Dahlberg (1963b) quoted Enoki's report that the frequency of fracture of the maxillary incisors was higher in European and American children who rarely had shovelling than in Japanese children with well-developed shovelling. However, as yet there do not seem to have been any biomechanical experiments on the structure of the anterior teeth.
Glancing at the structure of the incisors, it seems unlikely that well-developed marginal ridges, especially in the maxillary incisors, are suited to mastication as in the molar teeth, or to slicing or shearing as in the carnassial teeth of flesh-eating animals. To the present author, this structure seems to be for resisting the forces which strongly pull the incisor forward. That is, incisors with highly developed shovelling seem to function in biting off or pulling an object after holding it tightly like a vise. Or, because the maxillary incisors are somewhat inclined forward, the marginal ridges may simply resist the forward component of force generated by biting things with the anterior teeth, as Hylander (1978) showed. It is well known that Eskimos with highly developed shovelling not only eat hard food such as dried meat, frozen fish or bones but also use their teeth for various purposes as a third hand (Hylander, 1977). Such behavior must be associated with the development of shovelling.
Kikuchi (1954) suggested a relationship between shovelling and overjet of teeth based on the fact that, in modern Japanese, shovelling was less frequent in individuals with normal occlusion than in those with malocclusion. Sakai (1954) reported that the degree of incisor articulation was greater in individuals with shovel-shaped incisors than in those who did not have such teeth, and, at the same time, stated that generally in Mongoloids there were many individuals with stegodontia or overjet of maxillary anterior teeth. Pinto-Cisternas and Figueroa (1968) reported that in a Chilean population shovelling was significantly more frequent in individuals with stegodontia or hiato-dontia. On the other hand, individuals with labidodontia have been said to have highly-developed shovelling in the Japanese population (Sakai, 1954). Sakai (1954) and Suzuki and Sakai (1957) stated that the reason why there were more individuals with stego-dontia or overjet in Mongoloids than in other populations was not clear. However, such a phenomenon may be interpreted as follows on the basis of the fact that Eskimos with well-developed shovelling have a low frequency of prognathy (Hylander, 1977) in addition to the fact that most of the Jomon people, who were prehistoric hunter-gatherers in Japan, had labidodontia (Koganei, 1934; Suzuki, 1981). That is, highlydeveloped shovelling is a structure which originally functions only under the condition of labidodontia. In many recent Mongoloid populations, except the Eskimo and similar groups, the mandible was reduced in response to unknown demands, and, as a result, the mandibular incisors started to strike the marginal ridges of the maxillary incisors which protruded backwards on the lingual surface, resulting in the overjet of the maxil-lary anterior teeth or in stegodontia. Therefore, overjet or stegodontia is a secondary and temporary phenomenon, not a character which has any important adaptive meaning. On the other hand, shovelling is a character which resulted from the adaptation of using the anterior teeth as a vise with which things could be held and pulled forwards. This is the view of the present author. It may be strengthened by the fact that a phenom-enon which resembles shovelling in the human incisors is also observable in the incisors of herbivorous ungulates (, 1920).
Next, some problems regarding the lingual tubercle will be discussed. It was stated in the previous section (2.4.3.) that the broad incisor crown seemed to need the existence of the lingual tubercles. In the narrow incisors of females, the spines of the lingual tubercle, if any, often compose the basal part of the marginal ridges. A large lingual tubercle may not be required in the narrow incisors because of a type of mechanical strength related to the narrowness. This seems to satisfactorily explain the fact that there are no lingual tubercles in the narrow lateral incisors of Peking man (Weidenreich, 1937).
Robinson (1956) thought that the mesiodistal elongation of the incisal edge in the incisor crown was related to a cutting function and that the enlargement of the crown base in the direction perpendicular to the incisal edge was related to the available space and mechanical strength. He furthermore inferred that as the labial surface of a tooth crown was continuous with the labial surface of the root, the base of the crown was expanded lingualward to adjust to the labiolingually long form of the cross section of the root, inevitably resulting in a protuberance at the lingual base. However, although it is certain that the mesiodistal width at the dental neck is narrow in australopithecines and modern man, that width is considerable in Homo erectus and Neanderthals who had strongly-developed lingual tubercles. Therefore, the above interpretation by Robinson (1956) is questionable.
Adloff (1938) considered that the lingual tubercles of the maxillary incisors were developed by the mandibular teeth striking the lingual bases of the maxillary ones. He stated that the lingual tubercles of the incisors were homologous to the lingual cusps of the premolars (Adloff, 1927). This view may morphologically be correct, but these two structures seem completely different functionally. The maxillary incisors of seals are bicuspid teeth with well-developed lingual cusps like the human maxillary premolars (Kraus and Jordan, 1965). These teeth are used for firmly gripping things such as fish together with the single-cusped mandibular incisors with incisal edges. The maxillary incisors of bears are also bicuspid like the human maxillary premolars (Scott and Symons, 1974), and seem to have almost the same function as those of seals. If the function of the lingual tubercle was associated directly with the occlusion as asserted by Adloff (1938), it would not be strange that the human maxillary incisors also became bicuspid teeth as seen in seals and bears. However, no incisor with such a form has been known in the history of man. In the case of human teeth, therefore, Adioff's (1938) opinion is not acceptable. From the results stated above, to the present author the lingual tubercle seems to be a structure for supporting and strengthening the crown of a broader incisor.
The structure and the functional roles of shovelling and the lingual tubercle have been discussed in the paragraphs above. However, these problems should be examined further from various viewpoints, especially the biomechanical one. Tratman (1950) pointed out that the shovel-shaped incisors of the Mongoloid people had short and thick roots, and that the longitudinal axes of the crown and the root were not in line but crossed at an angle when observed from the mesial or distal side. It seems natural that the morphology of the root also changes owing to presence or absence of shovelling. Also, in non-human primates (Remane, 1960; Swindler, 1976) and other mammals, especially ungulates (, 1920), a structure like shovelling is observable. In order to clarify the connotation of such morphology, the way of using teeth, food habits, etc. must also be inspected in the future.
2.5. Summary
The implications of shovelling of the maxillary incisors were discussed through an analysis of the interrelations of shovelling and its component characters as well as with the mesiodistal crown diameter.
The relations between the characters of the maxillary central incisor were examined by the method of principal component analysis. This was performed using two different samples in order to assess the repeatability of results. The results show that the first principal component extracted had a high positive correlation with shovelling, the depth of the lingual fossa and the mesial and distal marginal ridges. This component was also correlated, though not so highly, with the mesial and distal spines of the lingual tubercle. These correlations with the spines were negative in males, but positive in females. The second principal component was correlated particularly with the mesio-distal crown diameter, the spines of the lingual tubercle and the central ridge.
Chiefly for interpreting the interrelations observed between shovelling and the spines of the lingual tubercle, the variation of these characters in modern and fossil hominids was surveyed through the literature. For the sake of convenience the human maxillary incisors were classified into three types. The first is an incisor with relatively weak shovelling and a narrow neck. This type is observable in australopithecines and most modern men. The second type has highly-developed mesial and distal marginal ridges running parallel in a wide crown, a deep lingual fossa and, at the same time, a well-developed lingual tubercle. This type is seen in Peking man, European Neanderthals, etc. The third type has highly-developed marginal ridges converging towards the neck and a less developed lingual tubercle. This type is observable in modern Eskimos and the like. From these variations, it was inferred that the between-sex difference in the contribution of the first principal component to the spines was referable to the betweensex difference in the mesiodistal crown diameter. In other words, male incisors with a broader crown tend to have spines flowing into the lingual fossa between the marginal ridges as in the case of Homo erectus or Neanderthals, whereas female incisors with a narrower crown tend to have spines which join the marginal ridges at the base of the tooth crown, resulting in a further development of shovelling. Shovelling itself seems to function relatively independently of the lingual tubercle. This was deduced from the fact that the first principal component which was strongly correlated with shovelling, and the second principal component which was highly correlated with the spines and the mesiodistal crown diameter were independent of one another, as well as from the state of shovelling and the lingual tubercle in the incisors of fossil hominids.
Finally, the problems to be solved in the future were pointed out through a review of previous reports on the structure and function of the incisors.