HOLOCENE OSTRACODS IN THE SOUTHERN BOSO PENINSULA
Paul M. Frydl
Mobil Oil Canada Ltd., Calgary, Canada
Conclusions and discussion
|
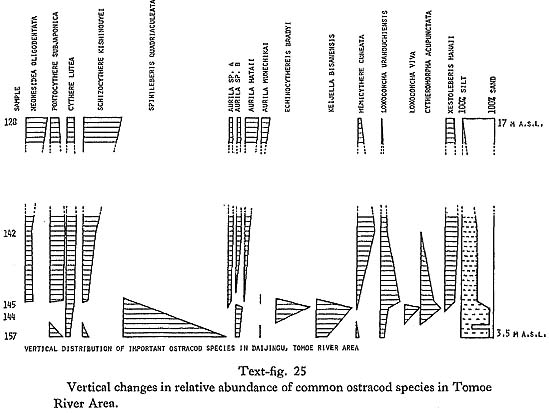
Interpretation of the past environment: I. Tomoe River Area: Changes in the relative abundance of the most common ostracods in sediments from 3 to 17 m above sea level in the lower part of Tomoe River valley are shown in text-fig. 25.
The lowest lying bluish green silty clay contains valves of Crassostrea gigas which is commonly found in shallow or intertidal near shore environments, often with lower salinity. The valves are not articulated but they show no signs of extensive transport and are probably not far from their place of origin. The ostracod assemblage (S sub-biofacies) is strongly dominated by Spinileberis quadriaculeata, a situation typical of innermost parts of bays with somewhat restricted circulation and some fresh water inflow. It is probable that silty clay was deposited in a brackish to marine environment, during the initial stages of the transgression, in a small incipient Tomoe Bay. Low sea level and proximity of shore is further indicated by the presence of peaty seams in the overlying sandy bed. The peaty sand bed is overlain by one meter of silt to sandy silt. Presence of articulated Ostrea denselamellosa, a marine oyster, at the base of the bed indicates deeper water and at the same time completely marine conditions. The ostracod assemblage from this bed belongs to the K sub-biofacies. It is characterized by an increase in Keijella bisanensis and Echinocythereis bradyi and accompanying decrease of Spinileberis quadriaculeata. These changes also indicate a change toward more marine conditions. The silty bed is abruptly overlain by a thin, organic matter rich bed. The lack of recognizable wood fragments, the nearly constant thickness of the bed, and its lateral continuity suggest that it is not a peat layer deposited during lowered sea level. It resembles the deposits of the dy biofacies (Ikeya and Hanai, 1980 MS) described from Hamana-ko estuary. Anoxic environment may develop in the inner parts of bays, where open-sea water circulation is not adequate, and which are too deep to be reached by wind generated waves. The presence of pyrite, indicating a reducing environment, as well as the absence of ostracods and other fossils, further supports this interpretation. The sediment overlying the organic rich bed is substantially different from the sediment below it. It consists of 50% sand size particles. There are changes in the ostracod assemblage, but they are not as abrupt as one might expect. Species characteristic of muddy bottoms of bays, which dominated the assemblages below the organic-matter-rich bed, decrease in abundance and eventually disappear. The phasing out of Spinileberis quadriaculeata, Echinocythereis bradyi, and Keijella bisanensis is matched by the increasing importance of sandy open-coast species: Neonesidea oligodentata, Pontocythere subjaponica, Schiziocythere kishinouyei, Aurila hataii, Aurila munechikai, and Hemicytherura cuneata. The sediment overlying the organic-matter-rich layer is characterized by great valiability; it consists of alternating beds of varying thickness and composition. An overall trend is apparent; with increasing elevation, the sediment increases in coarseness and becomes better sorted. Ostracods characteristic of the open coast increase in importance and the final assemblage is dominated by Schizocythere kishinouyei and Neonesidea oligodentata. This is probably due to continued sea level rise, which resulted in the widening of the bay mouth and filling in by sediment, which kept the bottom within the reach of wave generated currents. No C14 data are available for this section. Strong indications that the lowest exposed sediment was deposited in a brackish, shallow water environment during the initial stages of the transgression, together with a sea level of at least 18.5 m by 7,400 ( ± 80) yr B.P., would indicate that the basal bed was laid down very early-perhaps around 10,000 to 9,000 yr B.P. This is further supported by the dating of a nearby bed at 10 to 11 m above sea level, dated at 7,690 ( ± 190) yr B.P. Text-fig. 26 shows similar changes in ostracod fauna in the upper part of the valley. The lowest S sub-biofacies is absent, probably due to lack of exposure. The lowest silty bed (at 13.3 m above sea level) with abundant Dosinella penicillata was dated at 7,630 ( ± 70) yr B.P., making it contemporaneous with sediments at about 10 to 11 m above sea level in the lower part of the valley. The ostracod assemblage belongs to the K sub-biofacies and indicates a typical inner bay environment. Appearance and dominance of the assemblage by Loxoconcha viva and Nipponocythere bicarinata at about 15 m above sea level, indicate a change in conditions, probably brought about by a further rise in sea level, which broadened the bay and improved access of open-sea water. The faunal change is accompanied by increased proportion of sand, indicating a higher energy environment. The amount of sand continues to increase until it reaches 90% at 20 m above sea level. Keijella bisanensis, Loxoconcha viva, and Nipponocythere bicarinata are gradually replaced as the dominant species. The new assemblage is dominated by Schizocythere kishinouyei, Hemicytherura cuneata, Aurila hataii and other species typical of an open coast environment. Comparison with the highest assemblage of the previous section is interesting. Even though important, Schizocythere kishinouyei and Neonesidea oligodentata do not dominate the assemblage to the extent of the upper beds in the lower part of the valley. Continued presence of species typical of the KN and K sub-biofacies also indicates that conditions were intermediate between the open coast and bay, a proposition fully supported by the outline of the reconstructed bay.
Summary of the development of Tomoe River area: Incipient bay was formed in the lower part of the valley about 9,000 yr B.P. (text-fig. 27a). Continued rise of sea level caused the shifting of the S sub-biofacies into inner parts of Tomoe River valley and its tributaries, while the K sub-biofacies replaced the S sub-biofacies in the lower parts of the bay (text-fig. 27b). Increased depth together with the presence of a barrier (possibly a sand bar) impaired circulation in the deeper water in the lower part of the bay, giving rise to an anoxic environment. Further rise in sea level or removal of the barrier or both restored normal marine conditions in the lower part of the bay where the KN sub-biofacies developed (text-fig. 27c). At the same time, the S sub-biofacies shifted further landward and was replaced by K sub-biofacies. Further rise in sea level widened the bay, facilitating circulation of open-sea water as well as movement of waves and currents, resulting in coarser sediment. K sub-biofacies was replaced by KN sub-biofacies and the S and K sub-biofacies moved to the inner parts of small bays. Shallowing, brought about by sediment infilling and possibly by partial lowering of sea level (uplift?) resulted in the development of a shallow open coast environment and NS biofacies in all, except the innermost, parts of the bay (text-fig. 27d). Uplift at about 5,500 yr B.P. resulted in the formation of the Numa I terrace and ended sedimentation in the larger part of the area (text-figs. 27e and f).
II. Tateyama city area: The oldest dated material is Saxidomus purpuratus, boring in the siltstone underlying the oyster reef near Nishigo. S. purpuratus lives in depths ranging from intertidal to 40 m, but is most common at depths of about 20 m. Barnea wanilensis inornata, whose boreholes are found alongside those of Saxidomus purpuratus, is most common at shallower depths of about 4 m. At the above locality, its numbers are exceeded by those of S. purpuratus and it is therefore probable that water depth at this locality was already 10 to 20 m by 7,330 (±120) yr B.P. Pretostrea imbricata, which forms the oyster reef is known to live in depths ranging from 10 to 50 m, a fact that agrees well with the above conclusion. Ostracod assemblages found in the interstitial sediment in the reef belong to the LK biofacies (text-fig. 28). The lower parts of the reef, where Alveopora verrilliana is the only coral present, are characterized by slightly higher proportions of Spinileberis quadriaculeata and Nipponocythere bicarinata. The decrease in the proportion of these species, typical of muddy substrata, coincides with the appearance of diverse coral fauna, dated 6,400 to 6,300 yr B.P., suggesting that absence of fine sediment or perhaps sufficient elevation above the surrounding muddy bottom, due to the growth of the reef, was responsible for the increased coral diversity. Increased abundance of Ambocythere japonica, a species found in coral-bearing sediments in the West Coast Valley area, also coincides with the coral rich zone in the oyster reef. Xestoleberis hanaii, characteristic of agitated hard substrate environments, is another species that is abundant in the sediment throughout the reef.
The cessation of the oyster reef growth coincides with an abrupt change of ostracod fauna. The assemblage from the overlying silty sand belongs to the NL sub-biofacies and is characterized by dominance of Loxoconcha viva and Nipponocythere bicarinata. Spinileberis quadriaculeata, Cythere lutea, Pontocythere subjaponica, and Xestoleberis opalescenta also increase in abundance. Mutilus assimilis and Argilloecia lunata, common in the reef sediment, are lacking; Loxoconcha kattoi, Aurila sp. B and Xestoleberis hanaii are reduced to insignificant proportions. Parakrithella pseudadonta becomes most abundant in the fragmented shell bed directly overlying the oyster reef, but decreases in abundance in the above sediments. Inferred age of the fragmented shell bed coincides with the time of emergence of the Numa I terrace indicated in other areas. It is possible that it originated by erosion of landward positioned muddy sediments as a result of the sea level drop. Spondylus barbatus from the top of the reef was dated at 5,800 yr B.P., while Dosinella penicillata from the directly overlying sediment was dated at 4,490 yr B.P. The 1,300 year gap is consistent with the interpretation of the reef as a local bathymetric high, and 1,300 years were needed for the surrounding sediment level to reach the top of the reef. Most of the faunal changes can be explained by the absence of species which lived on the reef or on algae and sea grasses supported by it. The existence of NL sub-biofacies up to 4,440 yr B.P. indicates that water depth remained in the 10 to 20 m range. Younger sediments were, at this locality, removed by fluvial erosion. Only one C14 date is available for the surrounding sediments. It suggests that the NL sub-biofacies, which occupies the central part of the Tateyama area, was likely deposited during the time interval from 7,000-8,000 to 4,400 yr B.P. The PL biofacies, occurring in the western part of the area, appears to have been roughly contemporaneous with the sediments overlying the top of the reef; since its depth of depostion was probably 5 to 10 m, and its present location is 15 to 20 m above sea level, the bed overlying the oyster reef was probably deposited at depths of 15 to 30 m. This is in good agreement with the depth at which Loxoconcha viva and Nipponocythere bicarinata occur in the present Tateyama Bay. III. West Coast Valley Area: Silty sand infilling in boulder conglomerate in the lower part of Koyatsu valley was deposited 7,740 (±90) yr B.P. It contains ostracods characteristic of the AA sub-biofacies, indicating Zostera sea grass covered the bottom in the area where open coast to bay mouth environment prevailed. Because Crassostra gigas from conglomerate at 14 m above sea level in the upper part of the valley was dated at 7,840 (±110) yr B.P., the depth must have been at least 13 m. Changes in the relative abundance of ostracods in the coral-bearing sediments in the upper part of the valley are shown in text-fig. 29. The conglomerate, in addition to ostracods typical of the AL sub-biofacies, contains a high amount of Mutilus assimilis and Callistocythere undata, which are common in some samples of the IR biofacies in the present Tateyama Bay. It also contains a higher amount of Parakrithella pseudadonta and Cytherois zosterae and a lesser amount of Aurila sp. A and Loxoconcha japonica than other samples of the same sub-biofacies. This indicates a shallow subtidal to intertidal environment: an interpretation which agrees well with the presence of in situ Crassostrea gigas, a species commonly found from intertidal zone to depths of 5 m.
The coral-rich bed resting on the conglomerate contains ostracods typical of the AL biofacies. As opposed to sample 11 from the basal conglomerate, Aurila sp. A and Loxoconcha japonica become more abundant. This suggests a shallow, sea grass covered, silty sand bottom embayment. An increase in the amount of Spinileberis quadriaculeata in the sediments overlying the in situ corals (sample 9) can be explained either by a lowering of sea level, resulting in a greater proximity to a fresh water source, or by a rise of sea level, which would result in a lesser agitation and allow deposition of finer sediment. The increase in S, quadriaculeata is not accompanied by a corresponding increase in Mutilus assimilis nor Callistocythere undata, species which characterize the intertidal to shallow subtidal environment of sample 11. It is probable that the change was caused by a rise of sea level. The subsequent decrease in the amount of Spinileberis quadriaculeata present, coupled with the increased proportion of Xestoleberis hanaii and X. sagamiensis, together with the continued presence of large numbers of. Aurila sp. A and Loxoconcha japonica, might indicate further rise of sea level, which resulted in widening of the bay, better circulation of open-sea water, and development of intertidal rocky biofacies even along the inner margins of the bay. Shells from the top of the bed were dated at 6,720 (±60) yr B.P. While corals were growing in Koyatsu, sediment containing ostracods of the AA sub-biofacies was being deposited in the central part of the neighboring valley of Shiomi, covering a conglomerate dated at 7,230 (±70) yr B.P. Ostracods of the AA sub-biofacies are found also in the overing sediments, along with in situ Clementia papyracea, Paphia undulata, Panopea japonica, and Dosinella penicillata, which has been dated at 6,370 (±165) yr B.P. The molluscan and ostracod assemblages suggest 10 to 15 m deep bay with sufficient influx of open coast sea water. Corals also flourished in other bays-Numa, Kamisanakura-along the coast. All of them are found surrounded by sediment containing the AL ostracod sub-biofacies. Dated corals from Numa range from 6,160 (±120) yr B.P. to 5,365 (±170) yr B.P., and shells from the upper part of the coral bed at Kamisanakura were dated at 5,500 (±70) yr B.P. Ostracod assemblage from Numa is rich in Spinileberis quadriaculeata, suggesting more inner bay environment, which is consistent with its location in the inner part of paleo-Tateyama Bay and in the center of Numa valley. The Kamisanakura ostracod assemblage (sample 36) is characterized by near absence of Spinileberis quadriaculeata and abundance of Xestoleberis hanaii, Parakrithella pseudadonta, and Pseudoaurila japonica, suggesting a shallower environment or greater lateral proximity to it. To summarize, the transgressing sea reached at least 14 m above sea level by 7,840 (±110) yr B.P. It continued to rise and sometime before the formation of the Numa I terrace it reached at least 25 m, during which time molluscan borings in Koyatsu cliffs originated. Abundant hermatypic corals appeared slightly after 7,800 yr B.P. and continued to nourish at least until 5,500 yr B.P. Sea level reconstruction: Relative changes of sea level over the past 10,000 years have determined the outline of the coast of the southern Boso Peninsula. The shape of the coastline extended profound influence on near shore sediment distribution and salinity, which are the two major factors controlling distribution of corals and ostracods. Knowledge of the relative sea level position is therefore crucial for understanding changes of coral and ostracod biofacies; moreover, information on distribution and succession of coral and ostracod biofacies helps to explain the movement of sea level. The precision of reconstructed sea level depends on the precision of dating and on the certainty with which the position of the dated material for past sea level can be estimated. Under ideal conditions, where dated peat and marine sediment beds are available, sea level changes can be determined with a high degree of accuracy. Reconstruction of past sea level changes in the southern Boso Peninsula is facilitated by the relative abundance of C14 dated material from Holocene sediments. Unfortunately, most of the material comes from sediments whose depth of deposition can not be easily estimated. There are only two dated peat deposits and one dating of beach rock. The approximate depth of deposition can be partially inferred from the fossil assemblage and sedimentary structures, but the resolution (in most cases, 5 to 10 m) is inherently far too coarse to detect most changes of sea level, considering that the maximum change in the area did not exceed 30 m. The complications arising from the wide depth range of most potential depth indicators, such as marine fossils and bedding structures, are further aggravated by the sediments that were found in narrow bays, where environmental factors other than depth must have been often decisive. The information provided by terrace elevations also must be scrutinized carefully, becaue even though formed at the same time and at the same relative height of sea level, the rate of deposition and erosion may have been different in different localities, thus producing different elevations. Finally, most outcrops are located in channels of rivers and streams and are at least seasonally below the water table. Contamination of the dated material is the probable cause of several spurious dates. However, even in the absence of reliable precise indicators of sea level, minimum sea level curves for each area can be drawn based on the elevation of the highest available dated material in marine sediments for the given area and time. In most cases, the minimum sea level will lie several meters below the true sea level, but it gives the absolute minimum value for given time and place. The minimum sea level curves drawn for the three areas, West Coast Valley area, Tomoe River area, and Chikura area, resemble each other in shape (text-fig. 30). The curves rise rapidly and appear to reach a peak slightly before 6,000 yr B.P. No marine sediments dated younger than 5,480 (±110) yr B.P. are found above 20 m. Even though the curves have similar shapes, they differ in the absolute values of elevations; the curve for Chikura area is the highest (reaching 23 m); the curve for Tomoe River area is intermediate; and the curve for West Coast Valley area is the lowest, reaching only 19 m. Given the evidence of differential uplift during historical earthquakes, it is tempting to ascribe the differences to the cummulative effect of such events in the past. It is necessary to note, however, that there is an alternative explanation. The depositional surface is seldom horizontal and often has different elevations in different areas. The elevation of the depositional surface, and thus dated material, depends on the elevation of the underlying bedrock and on the rate of deposition. Thus, material of practically the same age is found at widely differing elevations in the upper and lower parts of Koyatsu valley. Similarly, the higher elevation of the Numa I surface in Chikura and Tomoe River areas in comparison with the West Coast Valley area could be due entirely to higher rate of deposition. The watershed of both Tomoe and Seto Rivers far exceeds that of the streams in the West Coast Valley area. Removal of sediments by waves and currents would also be slower in the inner parts of protected bays (such as Tomoe Bay) resulting in formation of a higher depositional surface. Accordingly, the variation in the elevation of terrace surfaces and material of the same age can be explained either by differential uplift or by differences in net deposition rate and location of dated samples. The presence of abundant bivalve borings up to 26 m in the Koyatsu valley and Tomoe River areas seems to indicate that the maximum sea level was the same in both areas, even though terrace surfaces are lower in the West Coast Valley area. It does not, however, prove conclusively that no differential uplift occurred. Uplift may have taken place, but evidence of sea level exceeding 26 m in Chikura area has either not been preserved or has not been found yet. In this sense there will always remain the possibility of discovering evidence of an even higher sea level in one of the areas, thus the question must be regarded as open. The simpler theory, involving no net differential uplift, has been provisionally adopted because it appears sufficient to explain nearly all the data available at the moment. The one exception is evidence of sea level at about 7 to 8 m above sea level around 7,240 (±80) yr B.P. in Maruyama area. The dated material comes from sediment which must have been deposited at a depth not exceeding a few meters. It is in direct contradiction with evidence from other areas, which indicates that sea level reached at least 21 to 22 m by this time. It is possible the material had been contaminated and thus yielded a younger age, but judging from the excellent preservation of the dated material and the low permeability of the sediment in which it was enclosed, contamination seems unlikely. The possibility of contamination cannot be rejected entirely, however, especially since the material comes from a river bank which is below the water table for at least part of the year. Lesser uplift of the area in comparison with the southern part of the area is another way to explain the anomalous date. The Maruyama area experienced the least uplift during the Taisho earthquake (1 m in comparison with the maximum of 2 m in Tomoe River area). The question cannot be resolved, however, until additional data from the area become available. Very little is known about the position of sea level prior to 8,000 yr B.P. The one clear indicator of a zero sea level, peaty seam at 4 m above sea level in Tomoe River area was not dated. Shells from 10 to 11 m above sea level from a nearby locality have yielded the age of 7,690 (±190) yr B.P. Assuming a similar rate of deposition as in the upper part of the Tomoe River valley, the age of the peat seam would be about 10,600 years. In Tomoe River area, Dosinella penicillata, living in 0 to 20 m depth and found in silt at 13.3 m above sea level, was dated at 7,630 (±70) yr B.P. The associated ostracod assemblage dominated by Keijella bisanensis indicates moderate depth. The overlying sediments containing ostracod species Keijella bisanensis and Nipponocythere bicarinata suggest a rise of sea level. Presence of the Keijella bisanensis assemblage in sediments further upstream at 18.5 and 20.5 m above sea level, which were dated at 7,400 (±80) and 6,530 (±80) yr B.P., confirms a continuing rise of sea level after 7,600 yr B.P. Crassostrea gigas from Koyatsu valley, dated at 7,840 (±110) yr B.P., is the oldest available date from the West Coast Valley and Tateyama areas. It lies at 14 m above sea level and indicates that sea level was at least that high by 7,840 yr B.P. C. gigas is commonly found in the intertidal zone but is present at a depth of several meters. Coral and ostracod fossil assemblages in sediment overlying the conglomerate with Crassostrea gigas suggest an increase in depth. Several dates, spanning about 2,000 years, for corals and shells from coral beds in the West Coast Valley area come from sediments about 17 m above sea level. The highest coral is from 19 m in Numa, dated at 6,160 (±120) yr B.P. The youngest of these dates was obtained from shells in coral bed in Uesanakura, located at 17 m above sea level. The ostracod assemblage dominated by Aurila sp. and Loxoconcha japonica, along with high coral diversity, suggests a depth of 5 to 10 m. Abundant bivalve borings were found in the cliff in Koyatsu valley and Tomoe River as high as 26 m, suggesting that the sea level reached at least that level and perhaps a few meters above it. This is consistent with the interpretation of the depositional environment of the Koyatsu coral bed by Hamada (1963), who concluded that the coral probably lived in a depth of about 10 m. The interval of highest sea level was not long enough to result in the creation of a wave-cut terrace. Crassostrea gigas from 21 m above sea level in Chikura area was dated at 7,620 (±110) yr B.P., and a shell sample from 22.5 m above sea level was dated at 6,910 (±200) yr B.P., indicating that sea reached at least this level by that time. C. gigas at 23 m above sea level yielded the age of 6,530 (±140) yr B.P. The C. gigas is overlain by peat at 23.5 m, dated at 6,200 (±135) yr B.P., suggesting that sea level was below 23 m at that time. We may say that level was between 23 and 23.5 m above sea level during the interval from 6,500 to 6,200 yr B.P. Wood contained in marine sediments lying near the surface of the Numa I terrace at 20 m above sea level in Mera, on the coast adjacent to the Tomoe River, yielded the age of 5,480 (±110) yr B.P., indicating that sea level remained above 20 m at least up to 5,480 yr B.P. If differential uplift has occurred, the presence of a peat bed in Maruyama area colld be explained by lesser uplift in the northeast: a hypothesis which would be also supported by evidence of lower sea level there around 7,240 (±80) yr B.P. If no differential uplift has occurred, the peat bed would provide evidence of an abrupt lowering sea level around 5,500-5,400 yr B.P. Presence of peat bed dated at 5,230 (±100) yr B.P. at about 18 m in Maruyama area indicates a lowering of sea level below 18m. The absence of sediments younger than 5,480 (±110) yr B.P. from Numa I terrace indicates a fall in sea level. The maximum extent of the lowering is uncertain. Nakata et al. (1980) suggested a lowering of sea level down to 14 m, based on unconformity they found in Tomoe River area (C14 dates TH 216 and TH 215). I was unable to locate and examine the outcrop of the unconformity. The brief description by Nakata et al. (1980) does not provide a strong indication of sea level lowered down to 14 m. The change in sediment type and erosion of underlying sediment (implied by the 1,000-year gap in the dates) could be accounted for by events taking place at depths of 8 to 10 m. The presence of shells dated at 4,740 (±100) yr B.P. and 4,280 (±130) yr B.P. in shallow sea sand (18 m above sea level) and in beach sand (18.9 m above sea level) in Chikura area suggests a new rise of sea level at least up to 18 m. Dated material close to the surface of the Numa II terrace indicates that it emerged around 4,300 yr B.P. The movement of sea level during the interval between the emergence of the Numa I and II terraces is not clear. Shells from cross-bedded sand, lying 0.3 m below the surface (at 10 m above sea level) of the Numa III terrace at Chikura were dated at 4,315 (±145) yr B.P. Shells from beach rock at the same elevation in a nearby locality yielded the age of 2,860 (±100) yr B.P. Cyclina sinensis from 10.3 m at Shirahama was dated at 3,360 (±100) yr B.P. Assuming that beach rock is the most precise indicator of sea level, the Numa III terrace emerged about 2,800 yr B.P. The sea level was at least 6 m high at 2,440 (±100) yr B.P., as indicated by dated shells from shallow marine sand in Heisaura. As recently as 820 (±80) yr B.P., the level was above 2.8 m in the same area (C14 date Gak 5459). The Numa IV terrace emerged during the 1703 earthquake. Two sea level curves for the southern Boso Peninsula have been published by Yokota (1978) and Nakata et al. (1980). The curves are in general agreement with each other and with the curve proposed in this study. Partly due to additional data and partly due to differences in interpretation the curve proposed here differs in several details (text-fig. 30).
New data have demonstrated that sea has reached high level about 2,000 years earlier than shown by Yokota. The wood from 4 to 5 m above sea level in Kawajiri River area is dated at 16,420 (±360) yr B.P. and lies in nonmarine sediments, which were probably deposited in the flood plain of Kawajiri River. It indicates only that sea level was below 4 to 5 m at that time. The curve proposed by Nakata et al. (1980) is appealing in its simplicity and emphasis on the main mechanism of sea level change in the area. There are, however, several points where the curve disagrees with available data and other points where there are no data to support the curve. For instance, the timing and extent of the drop of sea level at 6,100 yr B.P. is not convincingly demonstrated by the disparity in the TH 215 and TH 216 dates and is in clear contradiction with dates TH 086 and TH 085 from Numa, Gak 5478 date from Mera, and Gak 8602 date from Kotsuka, all of which demonstrate that sea level was still high at the time of its purported lowering to 14.5 m. Lowering of sea level is, however, indicated by the peat bed in Maruyama prior to 5,230 yr B.P. (if no differential uplift occurred), following which it appears to have risen in accordance with Nakata's curve. Further, there is no evidence of rise in sea level in the interval separating the emergence of Numa II and III terraces as shown in Nakata's curve. The sand from which sample TH 108 was obtained could have been deposited in a depth reaching several meters, but beach rock of sample Gak 3070 was most likely formed close or above the mean sea level. The curves of Yokota (1978) and Nakata et al. (1980), representing the minimum sea level curve, are correct only if all the dated material through which the curve passes were deposited close to sea level-a supposition which is clearly not true. The curves therefore underestimate the maximum elevation which the sea level reached. The curve proposed in this study suggests a sea level reaching at least 26 m. While supported by strong evidence of bivalve borings, the age and duration of this high stand of sea level has not been proven and therefore must be regarded with caution. Presently available data seem to favor the lowering of sea level in several steps in the interval between the emergence of the Numa I and II terraces. The scarcity of data does not allow a reconstruction with confidence of the movement of relative sea level after the emergence of Numa II terrace. Effect of the climatic change on ostracods: Climatic reconstructions based on fossil pollen indicate a warming up from around 9,000-8,000 yr B.P. and temperatures 1 to 2°C higher than at present in the interval from 8,000 to 4,900 yr B.P. (Kuroda and Hatanaka, 1979). The climatic optimum was reached around 6,000 yr B.P. The warm period was followed by an interval during which temperatures were several degrees lower than at present. Present-day climatic conditions extend back to about 1,500 yr B.P. (Yasuda, 1978). The above paleoclimatic reconstruction agrees well with the result of the study of Holocene molluscs in southern Kanto by Matsushima (1978). He found that temperate molluscs appeared around 9,500-8,700 yr B.P. and were followed by molluscs of the tropical type around 6,500-6,000 yr B.P. The latter disappeared around 4,000 yr B.P. The appearance and time of greatest diversity of hermatypic corals in the southern Boso Peninsula also coincides with the period of warm climate outlined above. While the appearance of both tropical molluscs and hermatypic corals is undoubtedly linked to the climatic amelioration, the connection between their disappearance or decreased diversity and the deterioration of climate is less clear cut. The suggestion by Matsushima (1979) that the disappearance of tropical molluscs was due to the disappearance of their habitat rather than due to climatic change deserves careful consideration. During the climatic optimum, the southern Boso Peninsula was characterized by a highly irregular coastline with drowned valley-type bays where hermatypic corals grew. The filling in of the bays, combined with tectonic uplift and eustatic lowering of sea level resulted in elimination of the drowned valley bay environment around 5,500-5,000 yr B.P. The effects of the climatic and physiographic changes on the corals are not easy to separate. However, because the present-day impoverished coral fauna of Tateyama Bay forms the northernmost limit of hermatypic corals in Japan, it seems unlikely that corals would have been able to survive the period of lower temperatures following the climatic optimum. Knowledge of the present Japanese ostracod distribution indicates that ostracod fauna is not as strongly affected by small variations in temperature as corals are. The slight warming (several degrees centigrade) around 6,000 yr B.P. would therefore be unlikely to produce a substantial change in the composition of ostracod assemblages. If the warmer climate had an effect, it would probably be the appearance of moderate amounts of few subtropical species, which are not presently found in the area. Examination of ostracod assemblages from fossil samples reveals that there is one species which might have appeared because of climatic change. Ambocythere japonica is present in most samples of the AL sub-biofacies and LK biofacies in amounts varying from 1 to 5%. All the samples come from sediments inferred to have been deposited during the climatic optimum and are associated with hermatypic corals. Ambocythere is a predominantly tropical genus; all but two occurrences reported come from the Caribbean and Southeast Pacific (one species was reported from Chile [Hartmann, 1963]; the other from Japan). In Japan, live Ambocythere japonica has been found only in Shikoku (Ishizaki, 1968). It is absent from samples taken in Hamanako estuary (Ikeya and Hanai, 1980 MS.), and in Tateyama Bay (this study); unpublished research indicates that it is also absent from Aburatsubo Bay in Miura Peninsula. Since the survey of recent ostracods in Japan is not complete and subsequent research may demonstrate presence of Ambocythere japonica in more northern latitudes, a definite conclusion can not be drawn at this point. The absence of A. japonica from other biofacies in sediments of the same age as those of the coral-bearing bed indicates that its presence is also controlled by environmental parameters other than temperature. However, its presence in two different biofacies in Jomon transgression sediments and its absence from Aburatsubo Bay, where the ostracod assemblage strongly resembles that of the AL sub-biofacies, warrant the speculation that the appearance and disappearance of A. japonica was the result of climatic changes. Effect of the transgression and regression on ostracod assemblages: The sea level changes during the past 10,000 years have produced environmental variations which are reflected in lateral shifts of ostracod biofacies. Even though the main cause of the variations (sea level change) was essentially the same throught the southern Boso Peninsula, the environmental change and the accompanying changes in ostracod biofacies were different in the different study areas. This is principally because topographic and bathymetric characteristics of an area can maximize or minimize the overall environmental change caused by sea level variation. Inspection of the outline of the present Tateyama Bay and that of the paleo-Tateyama Bay at the peak of the transgression demonstrates that the two are very similar. The same is true for a reconstructed bay at any sea level between 20 and 0 meters. The sediment distribution in the paleo-Tateyama Bay also agrees well with that of the present Tateyama Bay; the central part is formed by silt and silty sand, while sand predominates in the peripheral parts. Comparison between ostracod biofacies of the recent and paleo-Tateyama Bay also reveals good agreement; minor differences are ascribable to sampling and preservation bias and perhaps the gentler slope of the paleo-Tateyama Bay. Along the western coast, the present-day and Jomon transgression costlines and sediment differ strongly. The present coast is characterized by a nearly straight coastline and well-sorted coarse to fine sandy sediment. In contrast, at the peak of the Jomon transgression, the western coast was formed by drowned valley bays filling with poorly sorted silty sand. Thus, sea level change in the West Caost Valley area had a sub-stantially larger effect than in the Tateyama area. As the sea level rose, the bays reached farther inland, producing a greater number of different environments. Initially, the AA sub-biofacies occupied the small incipient bays, but as the sea level rose, a more quiet environment developed in the inner parts of the bay where the amount of agitation was small. AL sub-biofacies developed in this new environment. A continuing rise of sea level extended the bays farther inland where the inferred S sub-biofacies developed. As the sea level fell again, the environments of each biofacies disappeared in reverse order; the AA sub-biofacies was the last to disappear and still survives in some places (Z biofacies). The greatest changes took place in the Tomoe River area, primarily due to the length and gentle slope of Tomoe River valley and its tributaries and to the comparatively large discharge of Tomoe River. The large discharge resulted in the formation of S sub-biofacies in the initial stages of transgression when the embayment was still relatively small. With a rising sea level, the S sub-biofacies gradually kept moving inland to stay at the head of the bay. As the water depth and width of the bay mouth increased, biofacies constituted by assemblages that were more open coast in character moved into the bay. The maximum variety of biofacies was reached at the peak of the transgression, when the S, K, and K.N sub-biofacies and NS biofacies were all represented. The high rate of sedimentation, however, resulted in the shallowing of the bay and expansion of the open coast NS biofacies. The falling sea level accelerated this process and environments of the other biofacies gradually disappeared, the NS biofacies being the only one able to "retreat" with the receding sea. Lack of sampling points does not allow reconstruction of the biofacies changes in the Chikura and Maruyama areas. It is probable that the changes taking place in Chikura area were very similar to those in Tomoe River area, because the two have similar topography and sediment distribution. In the Maruyama area the influx of fresh water and fluvial sediment seem to have a much bigger role than in Tateyama area. |